Download presentation
Presentation is loading. Please wait.

1
35장 전사(RNA 합성) RNA 합성 가공 발현 조절
 RNA 합성 가공 발현 조절")
2
전사의 특성 유전자(gene): 여러 가지 다양한 RNA 종을 암호화하고 있는 DNA 구역
전령 RNA(messenger RNA, mRNA): 단백질 합성 정보를 암호화 전달 RNA(transfer RNA, tRNA): 아미노산을 리보솜으로 전달 리보솜 RNA(ribosomal RNA, rRNA): mRNA를 단백질로 번역하는 기구의 필수 요소로 기능 대장균에는 한 종류의 RNA 중합효소가 있지만, 포유동물 세포에서는 RNA 유형별로 서로 다른 종류의 RNA 중합효소가 합성에 관여
: 단백질 합성 정보를 암호화. 전달 RNA(transfer RNA, tRNA): 아미노산을 리보솜으로 전달. 리보솜 RNA(ribosomal RNA, rRNA): mRNA를 단백질로 번역하는 기구의 필수 요소로 기능. 대장균에는 한 종류의 RNA 중합효소가 있지만, 포유동물 세포에서는 RNA 유형별로 서로 다른 종류의 RNA 중합효소가 합성에 관여.")
3
1. 세포 내 RNA는 RNA 중합효소에 의해 합성된다
전사(transcription): DNA 주형에서 RNA를 합성하는 과정 모든 전사는 RNA 중합효소(polymerase)에 의해 촉매됨
: DNA 주형에서 RNA를 합성하는 과정. 모든 전사는 RNA 중합효소(polymerase)에 의해 촉매됨.")
4
RNA 중합효소의 필수 요소 1. 주형: DNA의 주형 가닥(template strand)의 염기 서열은 RNA 전사물의 염기서열과 상보적 DNA의 암호 가닥(coding strand)은 U 대신에 T가 있는 것만 제외하면 RNA 전사물과 동일한 염기 서열을 가진다 암호 가닥[의미 (+) 가닥, sense (+) strand], 주형 가닥[역의미 (-) 가닥, antisense (-) strand] 2. 활성화된 전구체: DNA 외에 필요한 기질로 RNA의 구성 성분인 ATP, GTP, UTP, CTP가 필요 3. 금속 이온: RNA 중합효소는 2가 양이온 (Mg2+ 또는 Mn2+) 보조인자를 필요로 함
은 U 대신에 T가 있는 것만 제외하면 RNA 전사물과 동일한 염기 서열을 가진다. 암호 가닥[의미 (+) 가닥, sense (+) strand], 주형 가닥[역의미 (-) 가닥, antisense (-) strand] 2. 활성화된 전구체: DNA 외에 필요한 기질로 RNA의 구성 성분인 ATP, GTP, UTP, CTP가 필요. 3. 금속 이온: RNA 중합효소는 2가 양이온 (Mg2+ 또는 Mn2+) 보조인자를 필요로 함.")
5
RNA 중합효소는 RNA 사슬의 개시와 연장을 촉매
- (RNA)n 잔기 + NTP (RNA)n+1 잔기 + Ppi RNA의 합성은 DNA의 합성과 유사 합성 방향이 5’ 3’ 사슬 연장 기전이 유사 pyrophosphate의 가수분해에 의해 합성이 추진 하지만, DNA 중합효소와 달리 RNA 중합효소는 시발물질 (primer)을 필요로 하지 않는다
n 잔기 + NTP (RNA)n+1 잔기 + Ppi. RNA의 합성은 DNA의 합성과 유사. 합성 방향이 5’ 3’ 사슬 연장 기전이 유사. pyrophosphate의 가수분해에 의해 합성이 추진. 하지만, DNA 중합효소와 달리 RNA 중합효소는 시발물질 (primer)을 필요로 하지 않는다.")
6
RNA 중합효소의 구조 대장균의 RNA 중합효소는 크고 (~500 kd) 다섯 종류의 소단위체로 이루어짐.
완전효소(holoenzyme): a2bb’ωs s 소단위체: 전사가 시작되는 부위를 찾아주고 RNA 합성의 개시에 참여한 후 효소의 나머지 부분들로부터 떨어져 나온다 이 소단위체가 없는 RNA 중합효소를 핵심효소(core enzyme)라고 부른다 (촉매자리를 포함)
: a2bb’ωs. s 소단위체: 전사가 시작되는 부위를 찾아주고 RNA 합성의 개시에 참여한 후 효소의 나머지 부분들로부터 떨어져 나온다. 이 소단위체가 없는 RNA 중합효소를 핵심효소(core enzyme)라고 부른다 (촉매자리를 포함)")
7
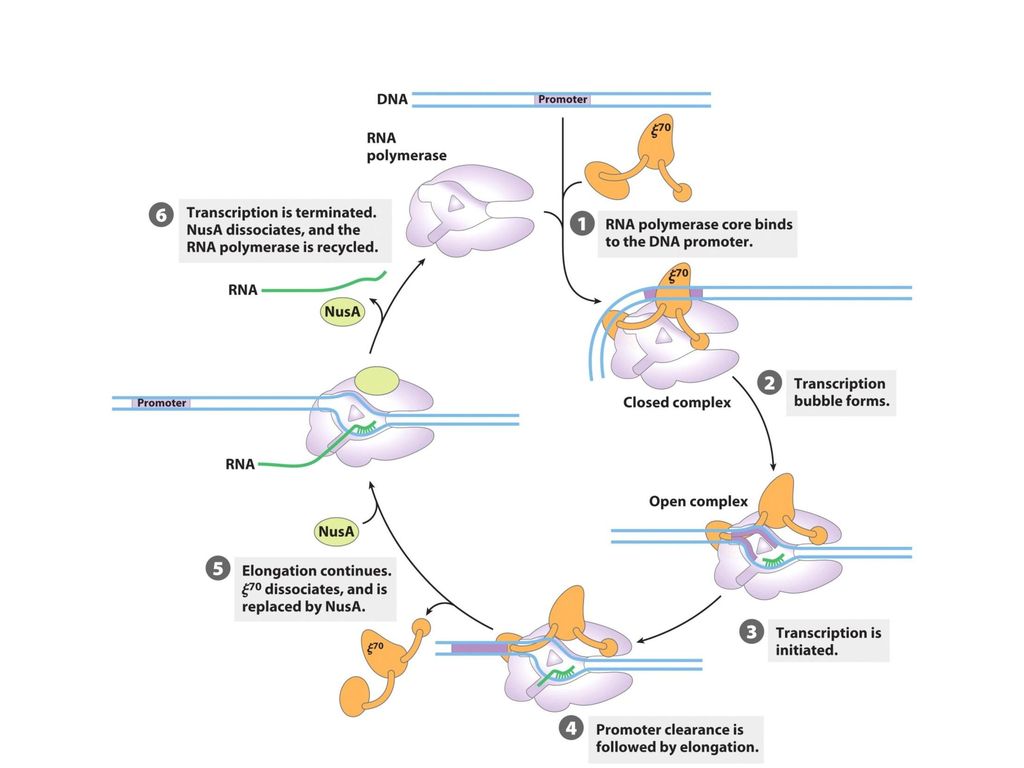
RNA 합성 과정: 1. 개시(initiation) 2. 연장(elongation) 3. 종결(termination)
 2. 연장(elongation) 3. 종결(termination)")
8
개시 촉진유전자(promoter)라고 불리는 특별한 DNA 염기 서열이 RNA 중합효소를 전사 개시 부위로 유도 전사 개시 부위의 5’ 위쪽에는 촉진유전자로 작용하는 두 개의 일치 염기 서열(consensus sequence) 존재; -10 (Pribnow box)과 -35 서열 -10과 -35 영역이 17 nucleotide(nt)만큼 떨어지는 것이 가장 적합 전사되는 DNA 염기 서열의 첫 번째 nt(개시 부위)는 +1로, 개시 부위 앞에 있는 nt는 -1로 표시 촉진유전자 서열의 효율성은 일치 염기서열과 유사한 정도에 따라 결정되어 전사를 조절한다 전사 인자(transcription factor, TF)는 촉진유전자 근처의 특정 서열에 결합하여 전사 효율에 영향을 준다 상류 요소(upstream element, UP 요소): 전사 개시 부위의 40~60 nt 정도 상류에 존재 UP 요소는 RNA 중합효소의 a 소단위체와 결합하여 RNA 중합효소가 결합할 수 있는 자리를 추가로 제공하여 전사의 효율을 증가시킴
 존재; -10 (Pribnow box)과 -35 서열. -10과 -35 영역이 17 nucleotide(nt)만큼 떨어지는 것이 가장 적합. 전사되는 DNA 염기 서열의 첫 번째 nt(개시 부위)는 +1로, 개시 부위 앞에 있는 nt는 -1로 표시. 촉진유전자 서열의 효율성은 일치 염기서열과 유사한 정도에 따라 결정되어 전사를 조절한다. 전사 인자(transcription factor, TF)는 촉진유전자 근처의 특정 서열에 결합하여 전사 효율에 영향을 준다. 상류 요소(upstream element, UP 요소): 전사 개시 부위의 40~60 nt 정도 상류에 존재. UP 요소는 RNA 중합효소의 a 소단위체와 결합하여 RNA 중합효소가 결합할 수 있는 자리를 추가로 제공하여 전사의 효율을 증가시킴.")
9
2) s 소단위체에 의한 promoter 인식 RNA 중합효소의 a2bb’ω 핵심만으로는 촉진유전자에서 전사를 시작할 수 없고 전사 개시가 제대로 일어나기 위해서는 RNA 중합효소가 정확한 개시 부위로 찾아가도록 s 소단위체가 필요 s 소단위체는 다른 일반 DNA 영역에 대한 RNA 중합효소의 친화도를 104배 정도 떨어뜨린다 친화도의 약화는 효소가 DNA 이중나선에 붙은 채 미끄러져 재빨리 촉진유전자를 찾게 해준다. s 소단위체가 없으면 핵심효소는 DNA에 무차별적으로 단단하게 결합 s 소단위체는 RNA 중합효소가 촉진유전자를 선택적으로 인식할 수 있게 해준다 DNA footprinting

10
s 소단위체는 새로운 RNA 사슬이 9~10개가 되면 떨어져 나온다. 그 다음 촉매 처럼 다시 핵심효소와 promoter에 결합
대장균에는 여러 유형의 촉진유전자를 인식할 수 있는 여러 종류의 s 소단위체가 있다. s 소단위체는 s70 (70 kd)라고도 불린다 다른 s 소단위체는 세균으로 하여금 온도 상승, 질소 고갈 및 그 외의 환경 변화에 적응할 수 있게 해준다
라고도 불린다. 다른 s 소단위체는 세균으로 하여금 온도 상승, 질소 고갈 및 그 외의 환경 변화에 적응할 수 있게 해준다.")
12
RNA 성장의 방향성 닫힌 촉진유전자 복합체(closed promoter complex)에서 17 bp의 DNA 구획이 열린 촉진유전자 복합체(open promoter complex)로 변하는 것은 전사에서 필수적인 과정이다. DNA 합성과 달리 RNA 합성은 시발물질 없이 바로 시작된다. RNA 사슬의 5’ 끝의 첫 염기는 pppG 혹은 pppA인데, 이는 RNA 사슬이 DNA 사슬처럼 5’3’ 방향으로 길어짐을 시사한다

13
2. 연장(elongation) RNA 합성의 연장 단계는 첫 phosphodiester 결합이 만들어진 후에 시작
σ 소단위체의 이탈은 핵심효소가 DNA 주형에 강하게 결합하여 전사거품(transcription bubble: RNA 중합효소, DNA와 갓 생긴 RNA 사슬이 존재하는 구역) 형성 개시 단계와 마찬가지로 연장 단계 전반에 걸쳐 약 17 bp의 DNA가 풀림. 갓 합성된 RNA 주형은 DNA 가닥과 약 8 bp의 혼성 나선(hybrid helix)를 만들고 혼성 나선에 있는 RNA의 3’-OH는 새로 들어오는 NTP의 a인산 원자를 공격
 형성. 개시 단계와 마찬가지로 연장 단계 전반에 걸쳐 약 17 bp의 DNA가 풀림. 갓 합성된 RNA 주형은 DNA 가닥과 약 8 bp의 혼성 나선(hybrid helix)를 만들고 혼성 나선에 있는 RNA의 3’-OH는 새로 들어오는 NTP의 a인산 원자를 공격.")
14
전사 거품은 1초에 170 Å (17 nm)의 속도로 이동 RNA 합성에서 낮은 정확도(1/104~105)는 여러 개의 전사물이 생성되고 전사의 오류가 자손에 전달되지 않기 때문에 허용 RNA 중합효소도 보조 단백질이 존재하면 교정 핵산분해효소 기능을 보인다

15
3. 종결(termination) phosphodiester 결합의 생성이 멈추고 RNA-DNA 혼성체가 분리된 후 DNA의 풀린 영역이 다시 감긴 다음 RNA 중합효소가 DNA에서 떨어짐 종결 신호 1) RNA 머리핀 구조 종결 신호는 전사되는 DNA 영역에 존재하지만 실제로는 종결 신호의 전사 산물이 전사를 종결시킴 DNA에 존재하는 종결 신호는 GC와 AT 풍부 영역의 자기 상보적 회문형(palindrome) 구조 RNA 중합효소는 머리핀으로 접히는 RNA 부분을 합성한 후 바로 전사를 멈춤 Oligo(U) 꼬리에서 만들어지는 RNA-DNA 혼성 나선은 불안정 약하게 결합된 RNA가 DNA 주형에서 떨어지고 RNA 중합효소도 떨어져 나온다 단일 가닥으로 된 DNA 주형은 재결합하여 이중 가닥 DNA를 다시 형성
 RNA 머리핀 구조. 종결 신호는 전사되는 DNA 영역에 존재하지만 실제로는 종결 신호의 전사 산물이 전사를 종결시킴. DNA에 존재하는 종결 신호는 GC와 AT 풍부 영역의 자기 상보적 회문형(palindrome) 구조. RNA 중합효소는 머리핀으로 접히는 RNA 부분을 합성한 후 바로 전사를 멈춤. Oligo(U) 꼬리에서 만들어지는 RNA-DNA 혼성 나선은 불안정. 약하게 결합된 RNA가 DNA 주형에서 떨어지고 RNA 중합효소도 떨어져 나온다. 단일 가닥으로 된 DNA 주형은 재결합하여 이중 가닥 DNA를 다시 형성.")
16
2) Rho 단백질 의존 전사 종결 머리핀-oligo(U)가 없는 전사를 종결시키기 위해서는 r라고 불리는 ATP 가수분해 능력을 가진 helicase 단백질이 필요 r 단백질은 단일 가닥으로 된 RNA의 72 nt 부분에 특이적으로 결합하고 RNA가 단백질의 중심을 통과하게끔 RNA와 결합 r의 ATP 가수분해효소 활성은 r로 하여금 RNA 중합효소를 따라서 RNA 사슬을 따라갈 수 있게 한다 r가 전사 거품에 있는 RNA 중합효소를 따라잡으면 RNA-DNA 혼성 나선을 분리시켜 전사 종결 단백질 의존성과 비의존성 종결의 공통점: 작용 신호가 DNA 주형이 아니라 새로이 합성되는 RNA에 있음

17
전사 과정 요약

18
진핵체의 전사 전사 개시 개시 복합체의 핵심: TATA 박스에 결합한 TATA 박스 결합 단백질 (TATA-box-binding protein, TBP) 안장 모양의 TBP (TFIID 복합체의 30 kd 성분)의 오목한 면에 소수성 상호작용으로 TATA 박스가 결합하여 DNA 이중나선을 풀고 작은 고랑을 확장시킴 다른 전사 인자들이 TBP의 안장 표면에 정해진 순서로 조립되어 기본 전사 기구 (basal transcription apparatus)를 형성 TBP가 결합한 후 순서대로 TFIIA, TFIIB, TFIIF, RNA 중합효소 II, TFIIE, TFIIH가 결합함
의 오목한 면에 소수성 상호작용으로 TATA 박스가 결합하여 DNA 이중나선을 풀고 작은 고랑을 확장시킴. 다른 전사 인자들이 TBP의 안장 표면에 정해진 순서로 조립되어 기본 전사 기구 (basal transcription apparatus)를 형성. TBP가 결합한 후 순서대로 TFIIA, TFIIB, TFIIF, RNA 중합효소 II, TFIIE, TFIIH가 결합함.")
19
RNA Pol II TFs

20
전사 연장 TFIIH에 의한 CTD의 인산화는 개시 단계로부터 연장 단계로 접어들게 만듦 인산화된 CTD의 역할 연장 과정의 안정화 연장 과정 중에 작동하는 RNA 가공 효소들의 유인

21
원핵과 진핵 생물의 유전정보 해석 다세포 진핵 생물은 모든 세포에 공통으로 존재하는 DNA의 전사를 차별적으로 조절함으로써 서로 다른 세포 유형을 만듦 유전자 발현에 영향을 미치는 진핵 생물 고유의 세 가지 특성 세 종류의 RNA 중합효소(RNA Pol I, II, III) 존재 Promoter 필요 다양한 종류의 promoter 요소의 존재와 조합 RNA 가공: 양쪽 끝의 변형, 일차 전사물의 이어 맞추기(splicing) 핵막 전사는 막으로 둘러싸인 핵에서, 번역은 핵 밖의 세포질에서 일어남 전사와 번역의 시간적, 공간적 분리 때문에 진핵 생물은 유전자 발현을 더 복잡하게 조절할 수 있음
 존재. Promoter 필요. 다양한 종류의 promoter 요소의 존재와 조합. RNA 가공: 양쪽 끝의 변형, 일차 전사물의 이어 맞추기(splicing) 핵막. 전사는 막으로 둘러싸인 핵에서, 번역은 핵 밖의 세포질에서 일어남. 전사와 번역의 시간적, 공간적 분리 때문에 진핵 생물은 유전자 발현을 더 복잡하게 조절할 수 있음.")
22
원핵 생물의 tRNA와 rRNA의 RNA 가공
원핵 생물에서 mRNA 분자들은 전사되면서 동시에 번역이 일어난다 tRNA와 rRNA 분자들은 갓 합성된 RNA 사슬의 절단과 변형을 통해 생성 하나의 일차 전사물에서 세 가지의 rRNA와 한 가지의 tRNA 분자가 잘려 나온다. 일차 전사물에는 간격(spacer) 영역이라 불리는 비암호화 영역도 존재. 다른 1차 전사물들은 여러 종류의 tRNA가 배열되거나 동일 tRNA가 여러 개 존재 붉은색: 메틸화, 푸른색: pseudouridine 화 검은색: dihydrouridine 화 1: RNase III 2: RNase P 3. RNase E
 영역이라 불리는 비암호화 영역도 존재. 다른 1차 전사물들은 여러 종류의 tRNA가 배열되거나 동일 tRNA가 여러 개 존재. 붉은색: 메틸화, 푸른색: pseudouridine 화. 검은색: dihydrouridine 화. 1: RNase III. 2: RNase P. 3. RNase E.")
23
1차 가공 ribonuclease P는 tRNA 분자의 올바른 5’ 말단을 만드는데 폴리펩티드 사슬 속에 촉매 활성을 가진 RNA 분자를 함유 (ribozyme) ribonuclease III: 이중나선 머리핀 영역의 특정 부분을 절단하여 5S, 16S, 23S rRNA 전구체를 만든다. 2차 가공: tRNA가 가공될 때 RNA 사슬 3’ 말단에 CCA 첨가. 3차 가공: RNA 분자의 염기와 리보오스가 변형된다 원핵생물에서 rRNA의 어떤 염기들은 메틸화된다 tRNA 분자는 효소에 의하여 uridylate가 전사 후 ribothymidylate과 pseudouridylate으로 변형

24
mRNA 가공 RNA 중합효소 II의 전사 산물이 가장 많이 변형되는데 대부분은 mRNA로 가공
RNA 중합효소 II로부터 막 생산된 산물은 선구 mRNA로 불리고 5’과 3’ 양 끝 모두 변형되며, 이어 맞추기(splicing)로 intron 제거
로 intron 제거.")
25
mRNA의 전사와 가공 연결 전사와 가공이 RNA 중합효소 II의 C 말단 영역(CTD, YSPTSPS 서열로 구성)에 의해 서로 통합 조절되는 증거가 있다. CTD의 2와 5번째 Ser의 인산화 상태는 여러 가지 kinase와 phosphatase에 의해 조절되고, RNA 전사와 가공에 참여하는 많은 단백질들과 결합할 수 있게 유도 RNA 중합효소 II의 활성은 CTD의 인산화에 의해 조절된다 CTD가 전사를 효율적으로 일어나도록 하기 위해 선구 mRNA로 유인되는 단백질들: 전사가 시작된 직후에 선구 mRNA의 5’ G을 메틸화하는 capping 효소 Intron이 합성될 때 각 intron의 절단을 촉발하는 이어 맞추기 기구의 성분들 전사물을 poly(A) 첨가 부위에서 자르고 3’ 아데닐화의 표적인 3’-OH기를 만드는 핵산 내부가수분해효소 이 과정들은 CTD의 인산화 상태에 따라 순차적으로 일어난다
 첨가 부위에서 자르고 3’ 아데닐화의 표적인 3’-OH기를 만드는 핵산 내부가수분해효소. 이 과정들은 CTD의 인산화 상태에 따라 순차적으로 일어난다.")
26
5’ 끝 가공(capping) 갓 합성된 RNA 사슬의 5’- triphosphate 끝은 전사 개시 직후에 변형
하나의 인산기가 가수분해되어 제거됨 5’-diphosphate 끝은 GTP 분자의 a-인산 원자를 공격하여 5’-5’ triphosphate 결합을 형성. 이를 cap이라 부름 끝에 위치한 G의 N-7 질소 원자는 S-adenosylmethionine에 의해 메틸화되어 cap 0가 됨 인접한 ribose들이 메틸화되어 cap 1과 cap 2를 만들기도 함 역할 cap은 탈인산화효소와 핵산분해효소로부터 5’ 끝을 보호함으로써 mRNA의 안정성을 높이고 진핵 생물의 단백질 합성 시스템에 의한 mRNA의 번역을 증진시킴

27
선구 mRNA의 이어 맞추기 (splicing)
고등 진핵생물의 대부분의 유전자들은 암호화 영역인 exon과 비암호화 영역인 intron (50-10,000 nt)으로 구성 이어 맞추기: intron은 제거되고 exon은 서로 연결되어 최종 mRNA가 만들어지는 과정 이어 맞추기 자리 intron은 GU로 시작되고 AG로 끝난다 척추동물에서 5’쪽 이어맞추기 자리의 일치 서열은 AGGUAAGU이고 GU는 변하지 않음 intron의 3’끝의 일치 서열은 10개의 pyrimidine(U 또는 C) 부분(polypyrimidine 구역)은 뒤에 어느 염기든 하나가 오고 그 다음 C가 오며, 끝에 반드시 AG가 온다 가지 자리(branch site): intron의 3’ 이어맞추기 자리의 위쪽으로 20~50 nt 사이 필수 요소: 5’과 3’쪽 이어 맞추기 자리, 가지 자리
으로 구성. 이어 맞추기: intron은 제거되고 exon은 서로 연결되어 최종 mRNA가 만들어지는 과정. 이어 맞추기 자리. intron은 GU로 시작되고 AG로 끝난다. 척추동물에서 5’쪽 이어맞추기 자리의 일치 서열은 AGGUAAGU이고 GU는 변하지 않음. intron의 3’끝의 일치 서열은 10개의 pyrimidine(U 또는 C) 부분(polypyrimidine 구역)은 뒤에 어느 염기든 하나가 오고 그 다음 C가 오며, 끝에 반드시 AG가 온다. 가지 자리(branch site): intron의 3’ 이어맞추기 자리의 위쪽으로 20~50 nt 사이. 필수 요소: 5’과 3’쪽 이어 맞추기 자리, 가지 자리.")
28
이어 맞추기 작용 방법 intron 내부의 A의 2’-OH가 intron 시작 부위의 GU의 phosphodiester 결합을 공격하여 새로운 2’-phosphodiester 결합을 형성하여 고리 모양의 intron을 형성한 후 절단된 exon 3’ 말단의 자유로운 3’-OH가 intron 3’ 말단의 AG의 phosphodiester 결합을 공격하여 intron 고리를 끊고 exon들을 연결함.

29
이어 맞추기 반응 U2와 U6가 intron 내부의 A가 intron의 5’끝의 GpU와 반응하도록 배열한다.
U5가 5’쪽 이어 맞추기 자리의 exon 서열과 상호 작용하고 곧 이어 3’쪽 exon과 상호작용을 시작하면서 이어 맞추기가 시작됨 U6가 U4에서 떨어지고 U2 및 intron의 5’ 끝과 상호 작용하여 U1을 spliceosome에서 떨어져 나가게 함 U2와 U6 snRNA가 spliceosome의 촉매 중심을 형성

30
3’ 끝 가공 대부분의 진핵생물 mRNA 분자들은 끝에 poly (A) 꼬리를 가지고 있음 이 꼬리는 전사가 끝난 후에 첨가
1차 전사물들은 성숙한 mRNA의 3’ 끝 너머로 수백 개의 nt를 가지고 있으며, poly (A) 꼬리가 첨가되기 전에 다 제거 선구 mRNA의 3’ 끝은 AAUAAA 서열을 인식하는 특정 핵산 내부가수분해효소(CPSF)에 의해 만들어짐 AAUAAA는 단지 절단 신호의 일부분이며 그 주위의 환경이 중요 선구 RNA가 절단되면, poly (A) 중합효소가 약 250개의 adenylate 잔기를 3’ 끝에 첨가 poly (A)의 역할 번역 과정의 효율과 mRNA의 안정성을 높인다는 증거들이 쌓임 70% mRNA 수명: 수 시간~일, 30% histone mRNA: <30 분
 꼬리가 첨가되기 전에 다 제거. 선구 mRNA의 3’ 끝은 AAUAAA 서열을 인식하는 특정 핵산 내부가수분해효소(CPSF)에 의해 만들어짐. AAUAAA는 단지 절단 신호의 일부분이며 그 주위의 환경이 중요. 선구 RNA가 절단되면, poly (A) 중합효소가 약 250개의 adenylate 잔기를 3’ 끝에 첨가. poly (A)의 역할. 번역 과정의 효율과 mRNA의 안정성을 높인다는 증거들이 쌓임. 70% mRNA 수명: 수 시간~일, 30% histone mRNA: <30 분.")
31
임상적 통찰 전사 저해 항생제 Rifampicin: RNA 합성의 개시를 특이적으로 억제. RNA 중합효소에 의해 만들어진 RNA-DNA 혼성체가 지나가야 할 통로를 막아서, RNA 사슬의 초기 phosphodiester 결합의 형성을 방해 actinomycin D: 이중나선 DNA에 강하고 특이적으로 결합. RNA 합성의 주형으로 사용되는 것을 방해 틈새 삽입(intercalation): DNA의 인접한 염기쌍 사이로 끼어들어감 원핵세포와 진핵세포 모두에서 RNA 합성을 억제할 수 있다. 항암제로 사용
: DNA의 인접한 염기쌍 사이로 끼어들어감. 원핵세포와 진핵세포 모두에서 RNA 합성을 억제할 수 있다. 항암제로 사용.")
32
임상적 통찰 대부분의 인간 선구 mRNA는 선택적 이어 맞추기 (alternative splicing)가 되어 한 개의 유전자로부터 여러 다른 단백질들을 만들어 낼 수 있다 선택적 이어 맞추기는 단백질의 다양성을 유발하는 보편적인 기전 동일한 유전자 내 exon의 여러 가지 조합은 특정한 조직이나 발생 단계, 혹은 신호 전달 경로에 필요한 독특한 형태의 단백질 형태를 만드는 성숙한 RNA로 이어맞추기 된다 이어 맞추기 자리의 선택: trans로 작용하는 이어 맞추기 인자와 선구 mRNA의 cis 작용 서열의 결합으로 결정 선택적 이어맞추기는 조합적 조절을 통해 유전체 서열의 융통성을 확장시키는 강력한 기전을 제공 이어맞추기의 결함으로 유발되는 여러 질병

33
원핵 생물의 유전자 발현 조절 특정한 환경의 변화에 적응하는 데 기여하는 일련의 효소들의 발현량은 함께 변하는데 유전자 발현의 동조 단위를 오페론(operon)이라 부른다 오페론은 조절 요소와 단백질을 암호화하고 있는 유전자들로 구성 b-galactosidase와permease, transacetylase의 동반 발현은 이 유전자들의 발현이 한 가지 방법에 의해 조절됨을 시사

34
락토오스 오페론(lac operon) 조절유전자(regulator gene): 억제자 유전자(i), 작동유전자(o), 촉진유전자(p)로 구성. 작동유전자 부위에 결합하는 lac 억제자 (repressor) 단백질을 암호화. 작동유전자(operator): 억제자가 결합하여 구조 유전자의 전사가 억제 3) 구조유전자(structural genes): z, y, a는 하나의 mRNA 분자로 전사. b- galactosidase (z), permease (y), transacetylase (a) 암호화 이처럼 두 개 이상의 단백질을 암호화하고 있는 mRNA 분자를 복수유전자(polygenic) 혹은 복수 시스트론(poly-cistronic) 전사물이라 한다
: 억제자가 결합하여 구조 유전자의 전사가 억제. 3) 구조유전자(structural genes): z, y, a는 하나의 mRNA 분자로 전사. b- galactosidase (z), permease (y), transacetylase (a) 암호화. 이처럼 두 개 이상의 단백질을 암호화하고 있는 mRNA 분자를 복수유전자(polygenic) 혹은 복수 시스트론(poly-cistronic) 전사물이라 한다.")
35
lac 억제자는 lactose가 없으면 작동유전자에 매우 강하게 결합하여 RNA 중합효소의 전사 작용 방해
Lactose에서 생성된 유도자(inducer)인 allolactose가 lac 억제자에 결합하게 되면 구조적 변화가 일어나 작동유전자 DNA에 대한 억제자의 친화도가 크게 감소하고 억제자-유도자 복합체는 DNA에서 떨어지고 오페론의 전사가 가능해짐. 유도자는 galactose와 포도당이 α-1,4 결합으로 연결된 lactose가 아니라 α-1,6 결합으로 연결된 allolactose임
인 allolactose가 lac 억제자에 결합하게 되면 구조적 변화가 일어나 작동유전자 DNA에 대한 억제자의 친화도가 크게 감소하고 억제자-유도자 복합체는 DNA에서 떨어지고 오페론의 전사가 가능해짐. 유도자는 galactose와 포도당이 α-1,4 결합으로 연결된 lactose가 아니라 α-1,6 결합으로 연결된 allolactose임.")
36
RNA 중합효소 접촉 단백질들의 전사 촉진 lac 억제자는 lactose가 존재할 때까지 전사를 억제
glucose가 고갈되면 lac 오페론의 전사를 촉진하는 서열 특이적 DNA 결합 단백질이 존재 cAMP 반응 단백질(cAMP response protein, CRP) 또는 대사산물 활성화 단백질(catabolic activator protein, CAP) Glucose의 농도가 감소되면 cAMP의 농도가 증가되어 CAP과 결합하고 lac 오페론이 활성화되어 전사 촉진 CAP-cAMP 복합체는 전사 개시 자리에서 -61 근처 부위에 결합 CAP가 붙어있는 촉진유전자로 RNA 중합효소를 유인하여 전사 개시를 약 50배 촉진시킴 포도당은 대사산물 억제(catabolic repression)라고 불리는 과정을 통해 lac 오페론의 발현을 억제
 또는 대사산물 활성화 단백질(catabolic activator protein, CAP) Glucose의 농도가 감소되면 cAMP의 농도가 증가되어 CAP과 결합하고 lac 오페론이 활성화되어 전사 촉진. CAP-cAMP 복합체는 전사 개시 자리에서 -61 근처 부위에 결합. CAP가 붙어있는 촉진유전자로 RNA 중합효소를 유인하여 전사 개시를 약 50배 촉진시킴. 포도당은 대사산물 억제(catabolic repression)라고 불리는 과정을 통해 lac 오페론의 발현을 억제.")
37
임상적 및 생물학적 통찰 박테리아 상호간의 신호전달에 의한 유전자 발현 조절
정족수 감지: Vibrio라는 오징어에 공생하는 박테리아간에 신호전달 물질 acyl homoserine lactone(AHL)을 분비하여 박테리아의 밀도가 일정수가 되면 luciferase라는 효소를 발현하여 생물 발광(bioluminescence) 시작 → 밤에 오징어의 발광 기관의 빛의 세기 약화하여 생존 도움 AHL은 세포의 수를 조절하는 자기유발물질임 다른 vibrio에 AHL과 결합하는 LuxR이라는 전사인자가 있어 luciferase의 효소를 생성하는 LuxA와 LuxB의 오페론의 promoter에 결합하여 전사 촉진 정족수 감지는 생체피막[biofilm, 치태(plaque)와 샤워찌끼 등]의 골격물질도 형성, 생체피막의 미생물은 면역 작용이나 항생제에 저항성 가짐.
을 분비하여 박테리아의 밀도가 일정수가 되면 luciferase라는 효소를 발현하여 생물 발광(bioluminescence) 시작 → 밤에 오징어의 발광 기관의 빛의 세기 약화하여 생존 도움. AHL은 세포의 수를 조절하는 자기유발물질임. 다른 vibrio에 AHL과 결합하는 LuxR이라는 전사인자가 있어 luciferase의 효소를 생성하는 LuxA와 LuxB의 오페론의 promoter에 결합하여 전사 촉진. 정족수 감지는 생체피막[biofilm, 치태(plaque)와 샤워찌끼 등]의 골격물질도 형성, 생체피막의 미생물은 면역 작용이나 항생제에 저항성 가짐.")
38
진핵체 전사 조절 높은 효율로 mRNA 합성이 일어나고 특정 유전자의 발현을 조직 선택적으로 촉진하려면 추가적으로 전사 인자들이 다른 부위에 결합해야만 함 매개 인자 (mediator)들을 유인하여 거대한 복합체를 형성하게 되고 이들이 기본 전사 기구와 상호작용하여 전사를 활성화 또는 억제시킴 매개 인자: 증진 유전자(enhancer)에 결합한 활성 인자들과 촉진유전자에 결합한 RNA 중합효소 II 간의 연결다리로 기능하는 25~30개의 소단위체로 이루어진 복합체
들을 유인하여 거대한 복합체를 형성하게 되고 이들이 기본 전사 기구와 상호작용하여 전사를 활성화 또는 억제시킴. 매개 인자: 증진 유전자(enhancer)에 결합한 활성 인자들과 촉진유전자에 결합한 RNA 중합효소 II 간의 연결다리로 기능하는 25~30개의 소단위체로 이루어진 복합체.")
39
증진유전자(enhancer) 수천 염기 쌍이 떨어진 거리에서 전사를 촉진함. 전사되는 유전자와의 상대적인 위치(상류, 내부, 하류; 같은 또는 다른 가닥)와 관계없이 효과를 발휘함 전사 조절에 참여하는 단백질과 결합함 여러 전사 인자로 인한 조절의 장점 조합적 조절(combinatorial control)이 가능 세포 내에 어떤 단백질들이 존재하느냐에 따라 조절 단백질의 기능이 달라짐 여러 가지 다양한 세포 유형을 가지고 있는 다세포 개체에게는 필수적 인간은 C. elegans 보다 2배 정도 더 많은 유전자를 가지고 있지만 조합적 조절에 의해 훨씬 더 많은 세포 유형을 가지고, 더 정교한 유전자 발현 조절이 가능함
이 가능. 세포 내에 어떤 단백질들이 존재하느냐에 따라 조절 단백질의 기능이 달라짐. 여러 가지 다양한 세포 유형을 가지고 있는 다세포 개체에게는 필수적. 인간은 C. elegans 보다 2배 정도 더 많은 유전자를 가지고 있지만 조합적 조절에 의해 훨씬 더 많은 세포 유형을 가지고, 더 정교한 유전자 발현 조절이 가능함.")
Similar presentations

? 초신성 폭발 원소들은.>")
 와 전자 (-) - 전기적 중성 - 원소의 종류마다 원자핵의 질량과 전자의 개수가 다름.>")










 (번역) 진핵세포의 경우.>")
 소재가 평면응력상태에 놓였을 때(σ3=0), 최대전단응력조건과 전단변형에너지 조건은σ1 – σ2 평면에서 각각 어떤 식으로 표시되는가? (2) σ1 =σ2인 등이축인장에서 σ = Kεn로 주어지는 재료의 네킹시 변형율을 구하라.>")
 유전학적으로 본 병인의 분류>")



