Download presentation
1
분자생물학 From DNA to Protein By 배형섭

2
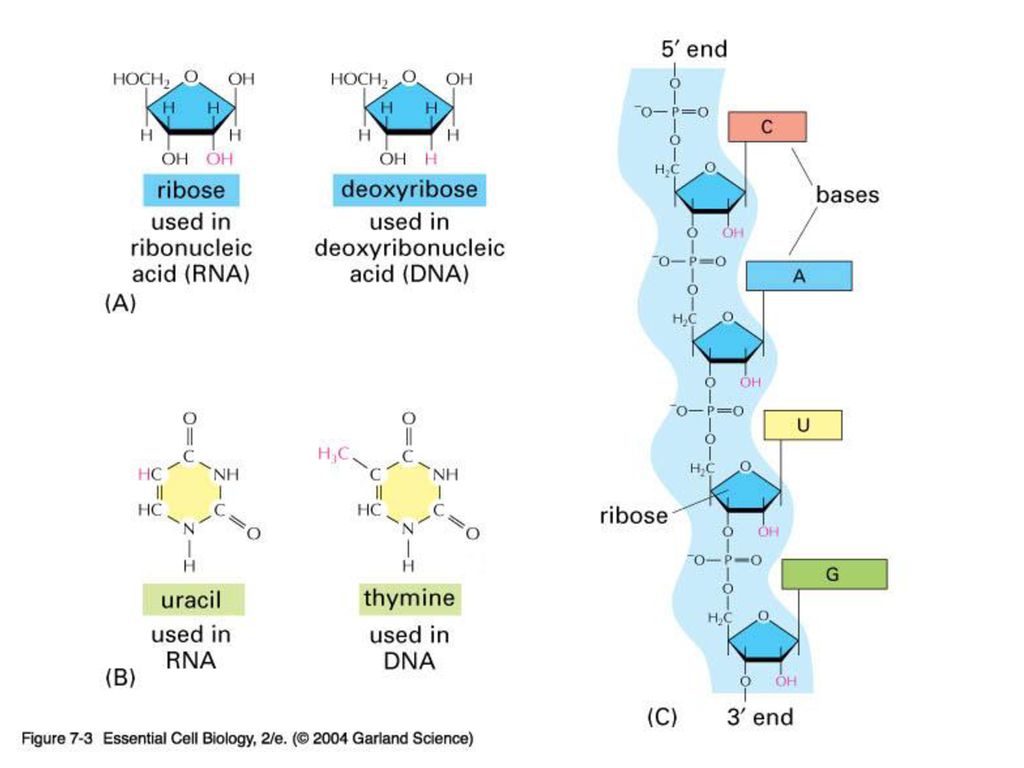
핵산의 구성 단위 핵산은 인산과 당을 포 함하고 있을 뿐만 아니 라 puerine계 2가지 화 합물
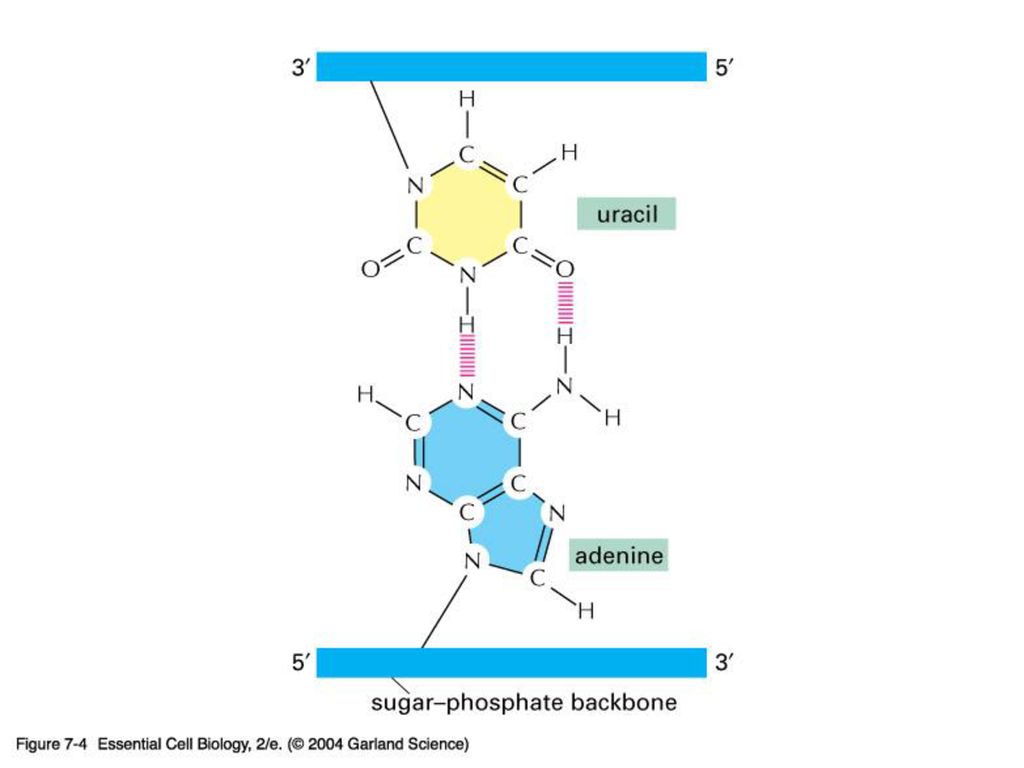
Adenine A, Guanine G pyrimidine계 Cytosine C, Thymine T, Uracil U(RNA의 경우) 을 포함한다.
 을 포함한다.")
3
Nucleotide DNA분자 내에서 인산 분자, 당 분자, 염기 하나가 모여 단위를 구성(이것들이 모여서 핵산을 이룬다)
")
4
DNA의 간단한 구조 왼쪽 그림에서와 같이 DNA는 염기의 수소결합에 의한 2중 나선 구조를 하고 있는데
AG의 이중결합과 CG의 삼중결합으로 붙어 있다.

5
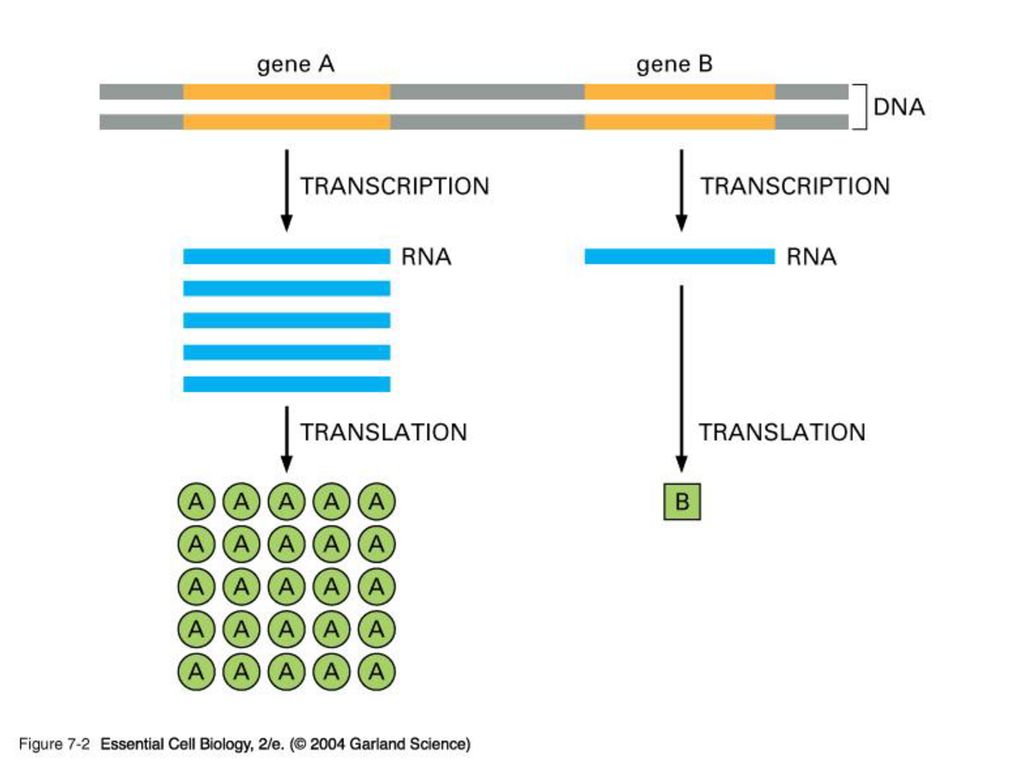
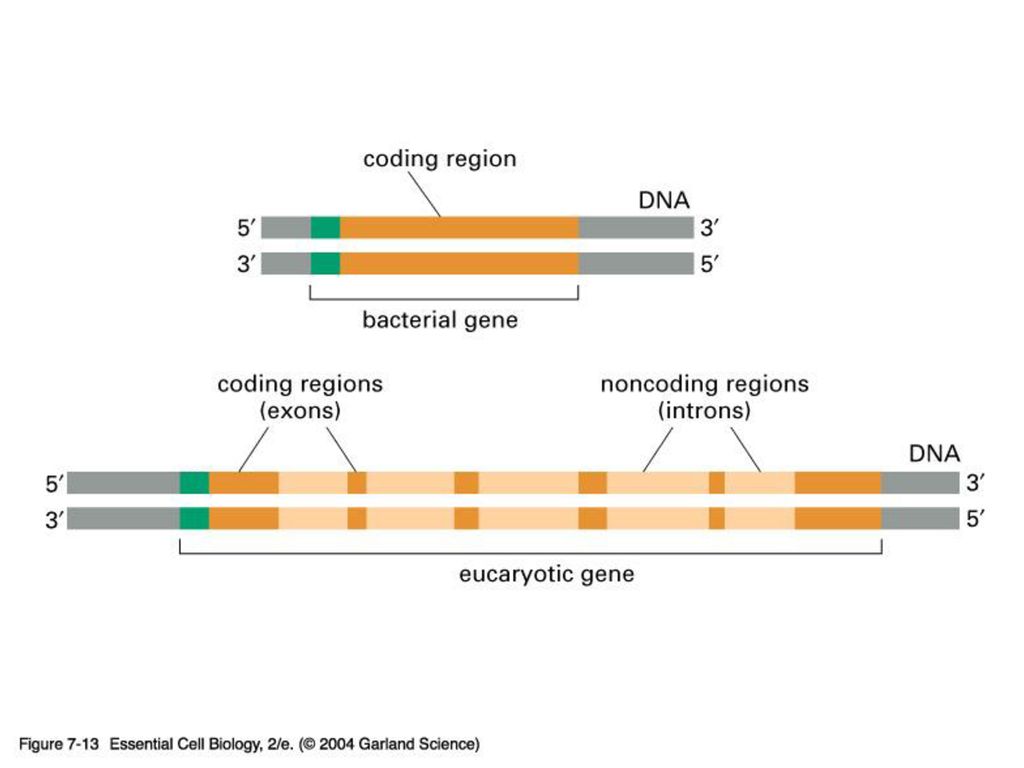
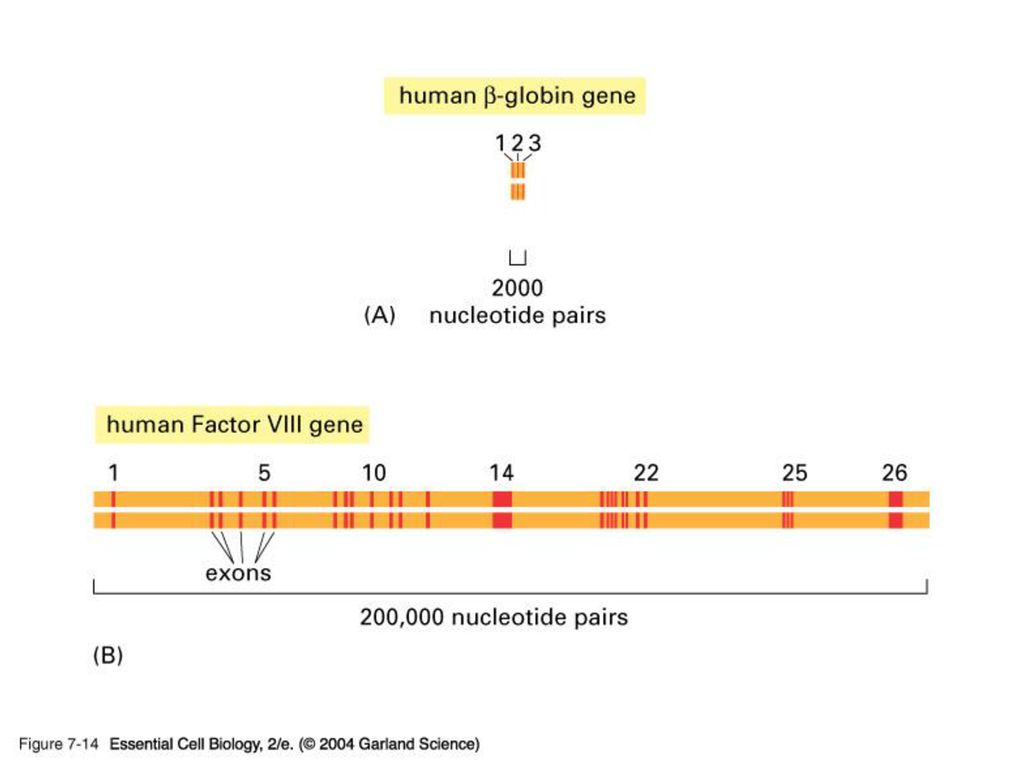
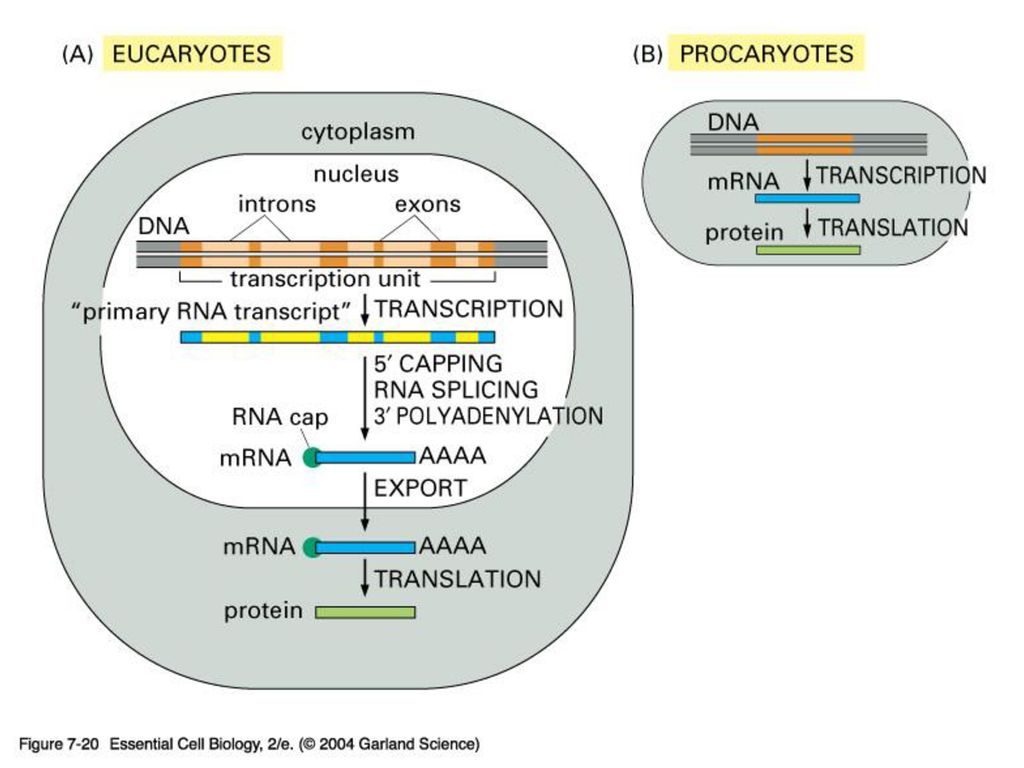
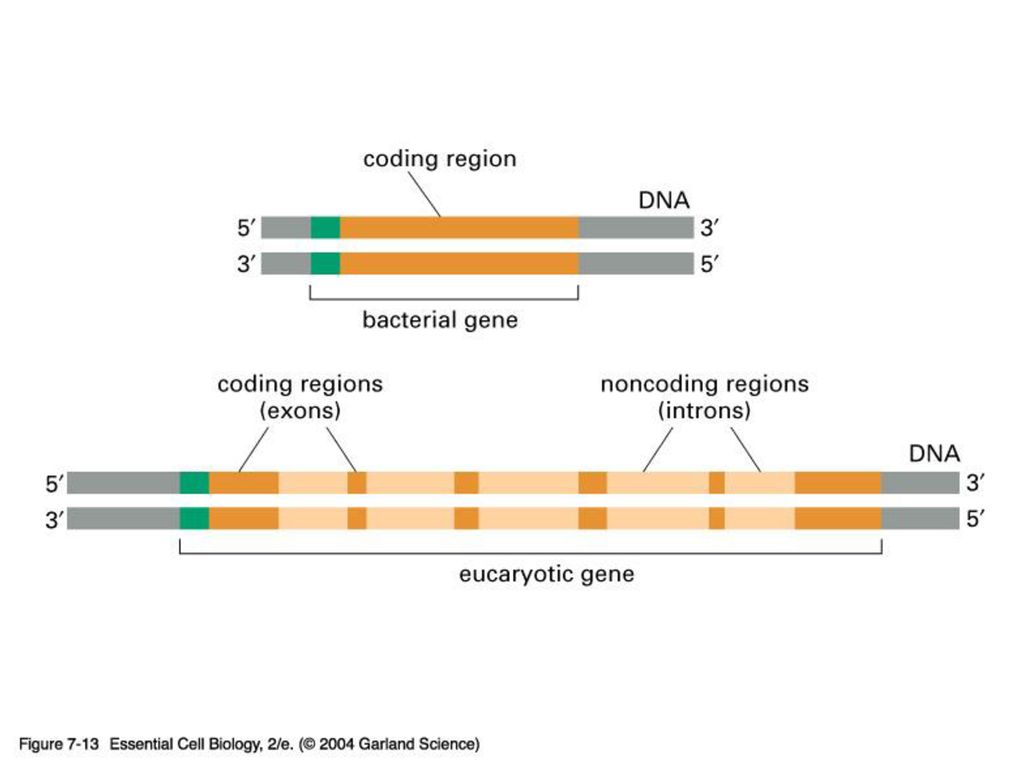
EXON & INTRON 원핵 생물처럼 핵이 없는 생물은 DNA의 유전암호가 전부 읽혀지는데 반해 포유동물 같은 진핵 생물은 유전 정보가 띄엄띄엄 읽혀지게 된다. 정의: intron은 암호화하지 않는 부분으로 그 유전자의 정보가 무엇이든 단백질을 합성하지 않는 유전자 서열이고 exon은 암호화하는 부분으로 그 유전자의 정보에 따라 단백질을 합성하는 유전자 서열임.

6
진핵 생물의 게놈에는 단백질을 만드는 유전자 서열에 intron이 기능이 없는 비암호화서열이 중간 중간에 끼여 있다
진핵 생물의 게놈에는 단백질을 만드는 유전자 서열에 intron이 기능이 없는 비암호화서열이 중간 중간에 끼여 있다. 즉 exon은 intron에 의해 몇 조각으로 나누어 져 있다. intron을 제거하고 exon을 이어 붙이는 과정을 splicing이라 함. 진화과정 중 intron을 경계로 하여 독특한 기능적 영역을 암호화하는 exon들이 다양하게 조합되어 새로운 기능을 보유하는 다양한 단백질을 지배하는 유전자들이 탄생되었다고 추정 중.

7
RNA

8
RNA의 종류(3가지) 1. mRNA(messenger RNA 전령 RNA): RNA polymerase에 의해
DNA(원본)의 한쪽을 풀어 그 염 기에 대응해 DNA의 유전정보를 충 실히 전달한다.(RNA에서는 T대신 U 가 대치된다.) DNA는 중요물질이기 때문에 핵 밖에 나갔다가 리소좀에 의해 분해 될 수 있기 때문에 전령인 RNA를 만들어 보내는 것이다. 이 과정을 전사(Transcription) 라 한다.
의 한쪽을 풀어 그 염. 기에 대응해 DNA의 유전정보를 충. 실히 전달한다.(RNA에서는 T대신 U. 가 대치된다.) DNA는 중요물질이기 때문에 핵. 밖에 나갔다가 리소좀에 의해 분해. 될 수 있기 때문에 전령인 RNA를. 만들어 보내는 것이다. 이 과정을 전사(Transcription) 라. 한다.")
9
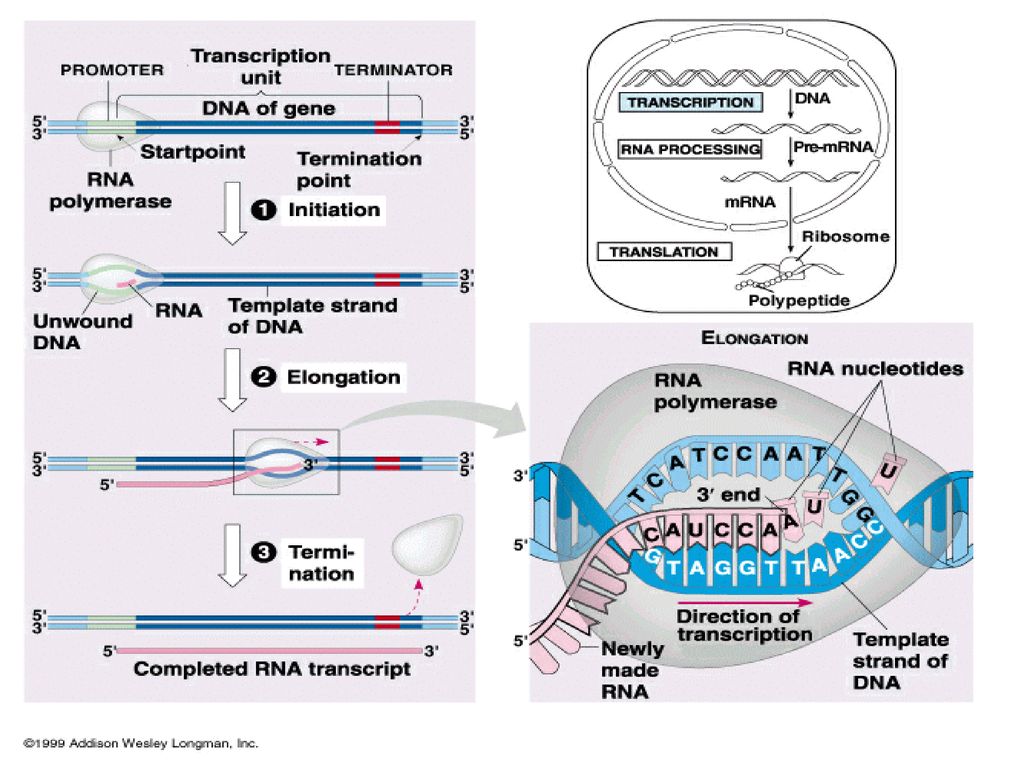
RNA 중합효소가 처음에 와서 붙는 위치를 프로모터라고 부른다. (promoter)
DNA복제나 RNA전사시 모두 5’말단->3’말단방향 으로 진행된다. (여기서 5’는 핵산의 당의 5번탄 소의 인산기이며 3’은 핵 산의 당의 3번탄소의 OH 기 이다)
")
11
2. tRNA(transfer RNA): 세포핵 내에서 합성된 mRNA는 핵 막의 핵공을 통해 세포질로 이동하여 리보솜에 부착하는데 이때 tRNA가 아미노산 특이성에 따라 특정 아미노산과 결합하여 리보솜에 부착되는 것이다. 간단히 말해 tRNA는 mRNA에서 온 염기서열에 대 응하는 아미노산으로 바꾸어 주고 이동시켜주는 통역 (translation)역할을 하는 것이 다.
역할을 하는 것이. 다.")
12
3. rRNA(ribosomal RNA) 단백질이 합성되는 세포 내 소기관인 리보솜(ribosome)을 구성하는 물질. 리보솜에서 아미노산들이 폴리펩타이드 결합으로 단백질이 된다. rRNA는 mRNA가 리보솜에 잘 붙을 수 있도록 도와주는 역할을 한다.

13
RNA를 거침으로써 얻어지 는 이익: DNA가 핵 밖으로 나갈 필요가 없다. (진핵 생물의 경우) 염기 서열의 일부만 발현된다. (발현 시기와 상대적 양 및 효율의 조절)
 염기 서열의 일부만 발현된다. (발현 시기와 상대적 양 및 효율의 조절)")
14
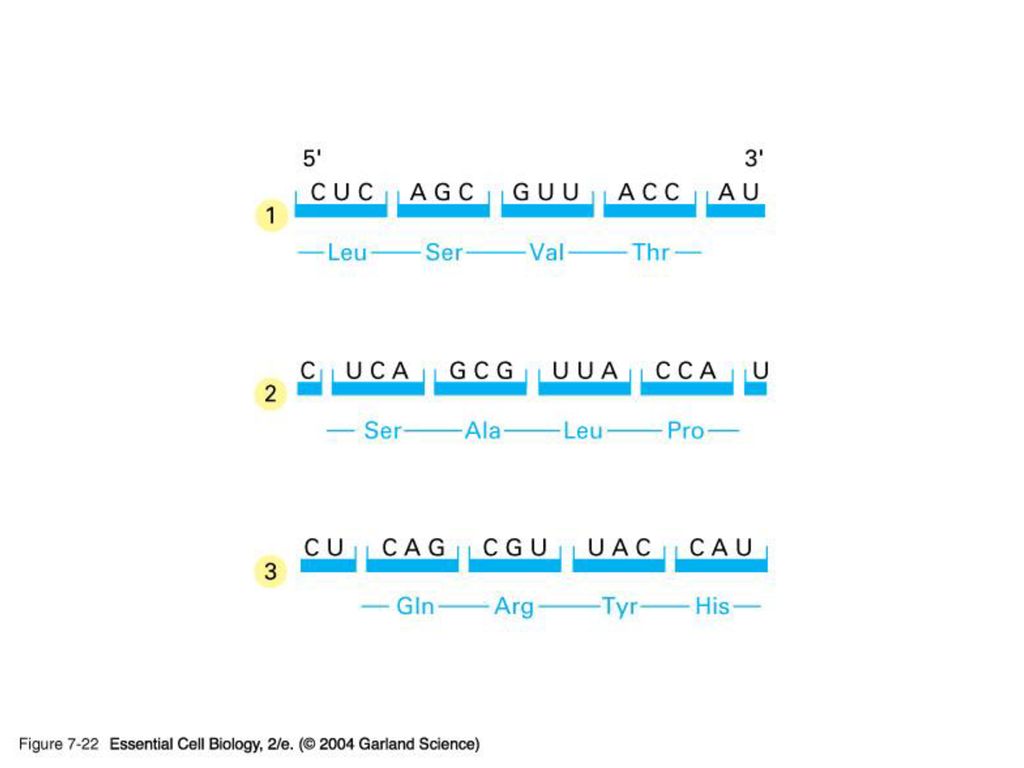
mRNA의 3조 암호문 염기 3개가 1개의 아미노산을 지정하며, 이를 코돈 (codon)이라 함
이라 함")
18
RNA는 단일 가닥으로 존재하기 때문에 다양한 방식으로 접혀서
독특한 3차 구조를 가진다. 이러한 성질 때문에 정보 저장 이외에도 여러 기능을 수행 할 수 있다. (=ribozyme.Splisosome이 대표적인 예)
")
19
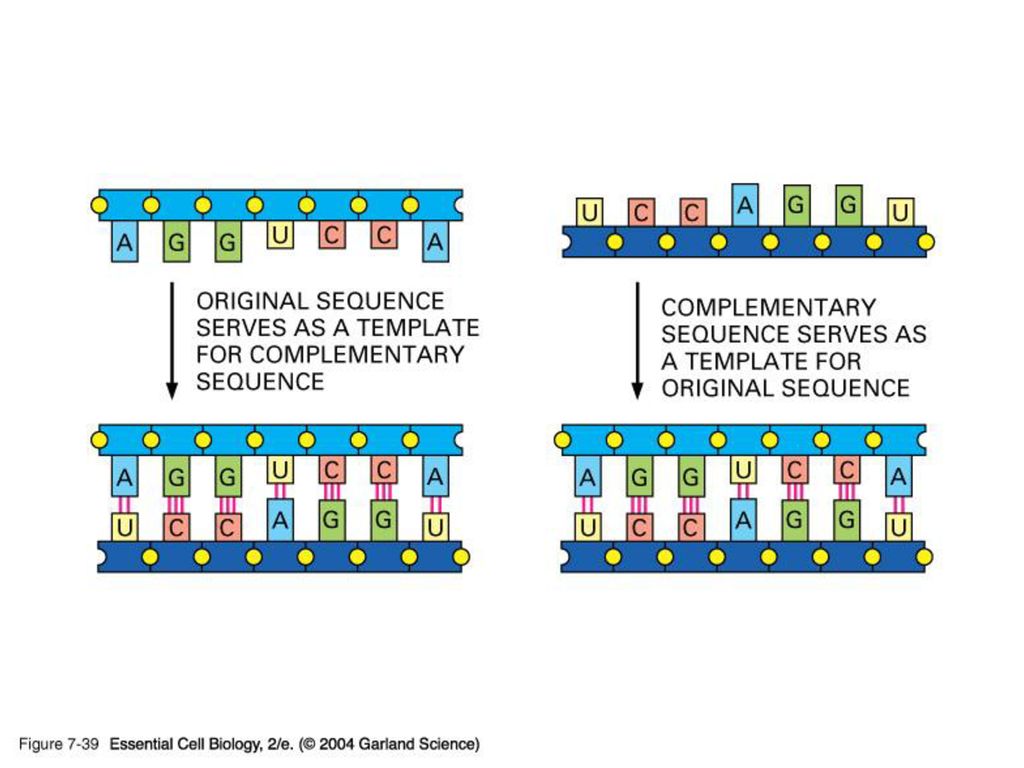
Transcription은 DNA 이중 나선 중 한 가닥 (=sense strand)을 주형으로 하여 일어난다.
염색체상에서 주형으로 쓰이는 가닥은 유전자마다 각기 다르다. ( 따라서 전사 방향 역시!) 실제로 만들어진 RNA 산물의 염기서열은 전사되지 않는 가닥(=antisense)과 동일하다. Antisense RNA/DNA 는 자연적 또는 인위적으로 세포내 유전자 발현 조절에 이용된다.
 실제로 만들어진 RNA 산물의 염기서열은 전사되지 않는 가닥(=antisense)과 동일하다. Antisense RNA/DNA 는 자연적 또는 인위적으로 세포내 유전자 발현 조절에 이용된다.")
20
RNA 중합효소는 DNA복제 떄 여러 다른 효소에 의하여 수행되던 기능을 모두 담당할 수 있다.

21
하나의 유전자에서 전사가 완결되기 전에 다시 앞쪽에서부터 전사가 시작되기도 한다.
활발하게 합성되는 종류의 단백질 유전자에서 특히 그러함. 아래 그림: 인접한 rRNA 유전자의 동시다발적 전사. (note) rRNA와 tRNA는 그 자체가 최종산물이며 단백질로 만들어지지 않음. (=so called Non-Coding RNA, 최근 rRNA와 tRNA 이외에도 많은 종류가 새롭게 발견됨. 핵과 세포질의 구별이 없는 원핵세포에서는 심지어 전사가 완결되기도 전에 그 시점까지 만들어진 RNA가닥에 ribosome들이 달라붙어 단백질 합성을 시작하는 경우도 있다.
 rRNA와 tRNA는 그 자체가 최종산물이며 단백질로 만들어지지 않음. (=so called Non-Coding RNA, 최근 rRNA와 tRNA 이외에도 많은 종류가 새롭게 발견됨. 핵과 세포질의 구별이 없는 원핵세포에서는 심지어 전사가 완결되기도 전에 그 시점까지 만들어진 RNA가닥에 ribosome들이 달라붙어 단백질 합성을 시작하는 경우도 있다.")
23
Transcription

24
Similarity between replication and transcription
Both processes use DNA as the template Phophodiester bonds are formed in both cases. Both synthesis directions are from 5 to 3

25
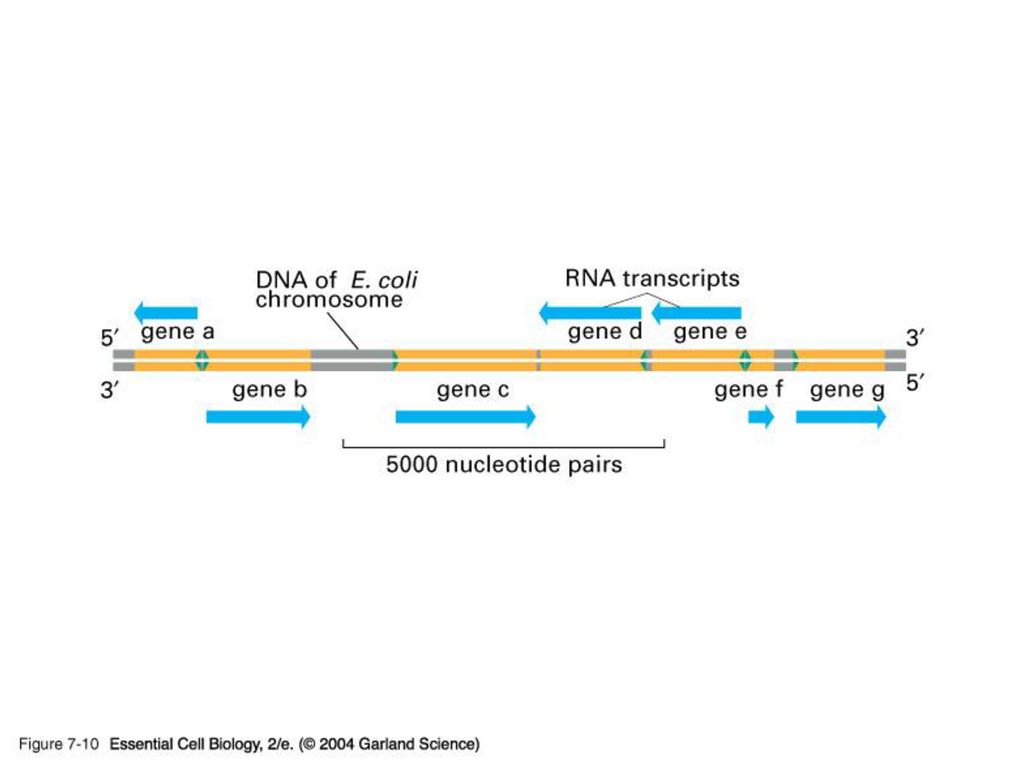
Template and Enzyme The whole genome of DNA needs to be replicated, but only small portion of genome is transcribed in response to the development requirement, physiological need and environmental changes. DNA regions that can be transcribed into RNA are called structural genes

26
Template The template strand is the strand from which the RNA is actually transcribed. It is also termed as antisense strand. The coding strand is the strand whose base sequence specifies the amino acid sequence of the encoded protein. Therefore, it is also called as sense strand

27
Asymmetric transcription
Only the tempate strand is used for the transcription, but the coding strand is not. Both strands can be used as the templates. The transcription direction on different strands is opposite. This feature is referred to as the asymmetric transcription.

28
Transcription process
RNA polymerase (an enzyme) attaches to DNA at a special sequence that serves as a “start signal”. The DNA strands are separated and one strand serves as a template. The RNA bases attach to the complementary DNA template, thus synthesizing mRNA. The RNA polymerase recognizes a termination site on the DNA molecule and releases the new mRNA molecule.
 attaches to. DNA at a special sequence that serves as a start signal . The DNA strands are separated and one strand serves. as a template. The RNA bases attach to the complementary DNA. template, thus synthesizing mRNA. The RNA polymerase recognizes a termination site on the. DNA molecule and releases the new mRNA molecule.")
29
Eukaryotic Transcription

30
Stages of Transcription -Initiation

31
Initiation Stayes put for a bit
• Nucleoside triphospates (NTPs) are added one by one until 9 bp have been added, and then elongation can occur (i.e. RNApoly can move)
 are added one by one. until 9 bp have been added, and then elongation can. occur (i.e. RNApoly can move)")
32
RNA Polymerase The enzyme responsible for the RNA synthesis
is DNA-dependent RNA polymerase. The prokaryotic RNA polymerase is a multiplesubunit protein of ~480kD. Eukaryotic systems have three kinds of RNA polymerases, each of which is a multiple-subunit protein and responsible for transcription of different RNAs.

33
RNA-pol of Prokaryotes: Holoenzyme
The holoenzyme of RNA-pol in E.coli consists of 5 different subunits:

34
Rifampicin, a therapeutic drug for tuberculosis treatment, can bind specifically to the subunit of RNA-pol, and inhibit the RNA synthesis. • RNA-pol of other prokaryotic systems is similar to that of E. coli in structure and functions

35
RNA-pol of eukaryotes RNA-pol I II III Products 45S Rrna hnRNA 5SrRNA
tRNA snRNA Sensitivity To Amanitin No high moderate Amanitin is a specific inhibitor of RNA-pol.

36
Three RNA polymerases One for each major type of RNA RNApoly I makes pre-rRNA RNApoly II makes pre-mRNA RNApoly III makes pre-tRNA Each polymerase has a different promoter structure

37
RNApoly II promoter Initiator sequence surrounding the start point
TATA box at about –25 bp TATA+Initiator = core promoter A transcription factor binds to the TATA box before RNAPoly II

38
Bacterial Promoters A promoter is where RNApoly binds
Template strand versus Coding strand Upstream – in the 5’ direction on the coding strand Consensus sequences

39
Transcription factors
A basal transcription factor is always required to allow RNApoly to bind to DNA For RNApoly II, TFIID binds to the TATA box. This is the basal transcription factor. More TFs bind to TFIID through protein-protein interactions to form the preinitiation complex. Then RNApoly binds Many TFs may be involved

40
Recognition of Origins
Each transcriptable region is called operon. One operon includes several structural genes and upstream regulatory sequences (or regulatory regions). The promoter is the DNA sequence that RNA-pol can bind. It is the key point for the transcription control.
. The promoter is the DNA sequence that RNA-pol can bind. It is the key point for the transcription control.")
41
Promoter regulatory sequences structural gene

42
Prokaryotic promoter Promoter RNA 5’ 3’
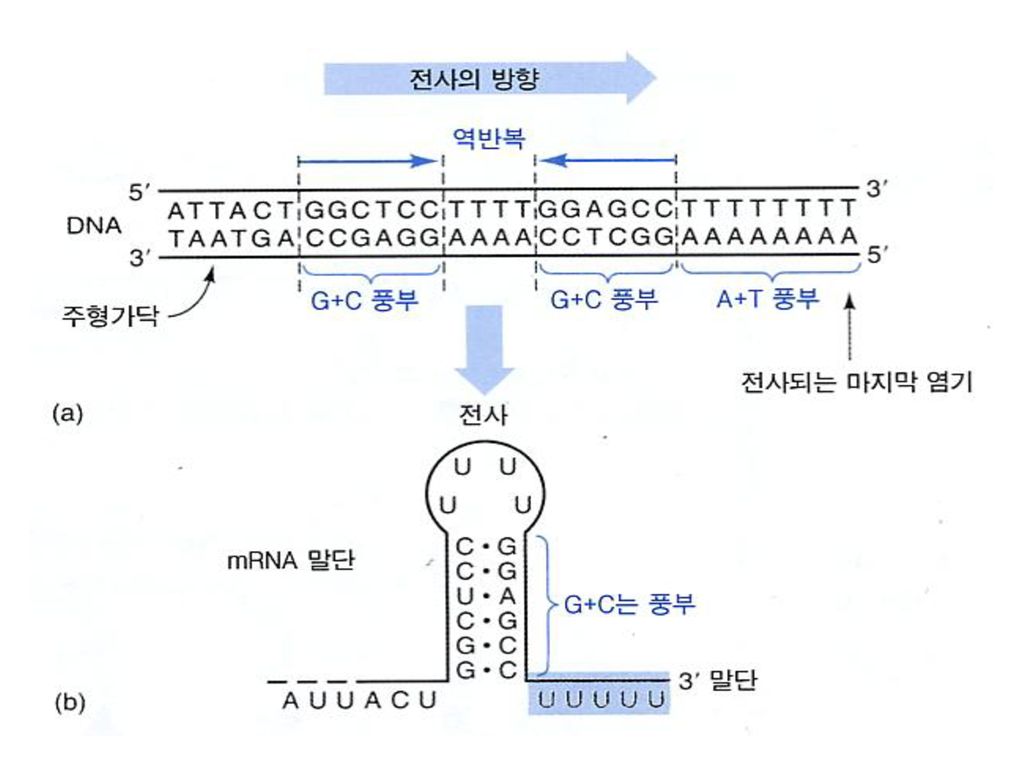
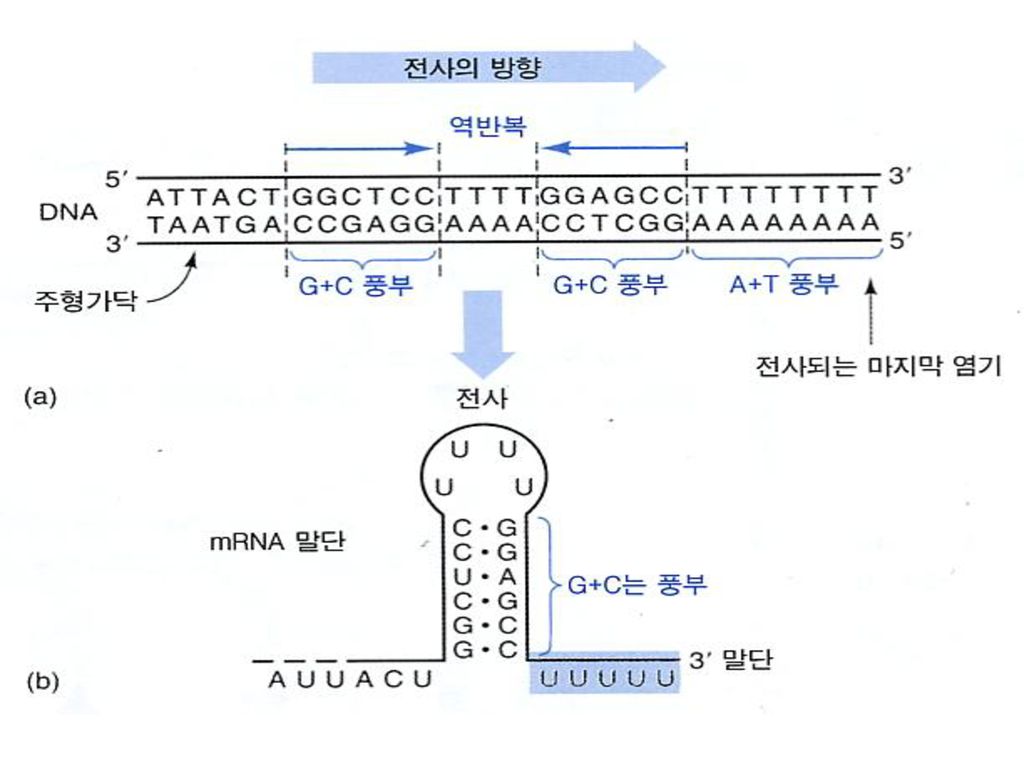
Regions of similarity are found around 10 and 35 bases before the start site of transcription: DNAse protection shows that RNA polymerase can bind to these same regions. Mutations of these sites can lead to the elimination or reduction of transcriptional initiation at a promoter. Differences in these sites control the relative rates of expression of different genes. Strong promoters have sites that are very similar to the consensus sequence while weak promoters show many differences

43
Consensus Sequence

44
The -35 region of TTGACA sequence is the recognition site and the binding site of RNA-pol.
The -10 region of TATAAT is the region at which a stable complex of DNA and RNApol is formed.

45
Transcription Process

46
General concepts Three phases: initiation, elongation, and
termination. The prokaryotic RNA-pol can bind to the DNA template directly in the transcription process. The eukaryotic RNA-pol requires cofactors to bind to the DNA template together in the transcription process

47
Transcription of Prokaryotes
Initiation phase: RNA-pol recognizes the promoter and starts the transcription. Elongation phase: the RNA strand is continuously growing. Termination phase: the RNA-pol stops synthesis and the nascent RNA is separated from the DNA template.

48
a. Initiation RNA-pol recognizes the TTGACA region,
and slides to the TATAAT region, then opens the DNA duplex. The unwound region is about 171 bp

49
The first nucleotide on RNA transcript is always purine triphosphate
The first nucleotide on RNA transcript is always purine triphosphate. GTP is more often than ATP. The pppGpN-OH structure remains on the RNA transcript until the RNA synthesis is completed. The three molecules form a transcription initiation complex.

50
No primer is needed for RNA synthesis.
The subunit falls off from the RNA-pol once the first 3,5phosphodiester bond is formed. The core enzyme moves along the DNA template to enter the elongation phase.

51
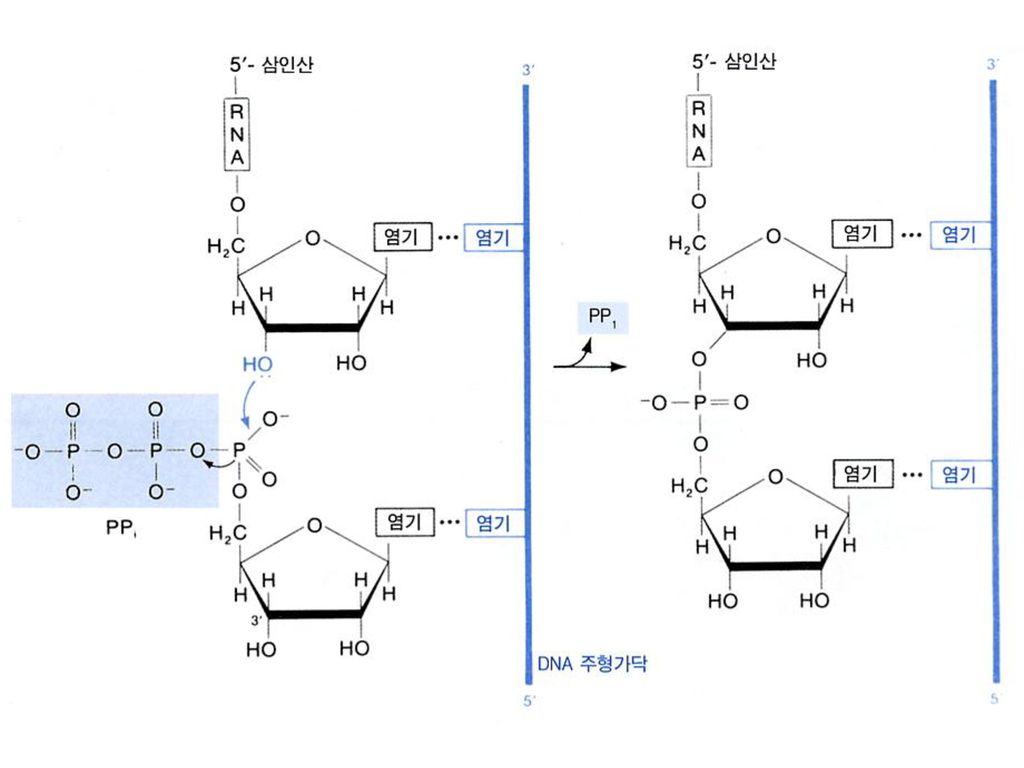
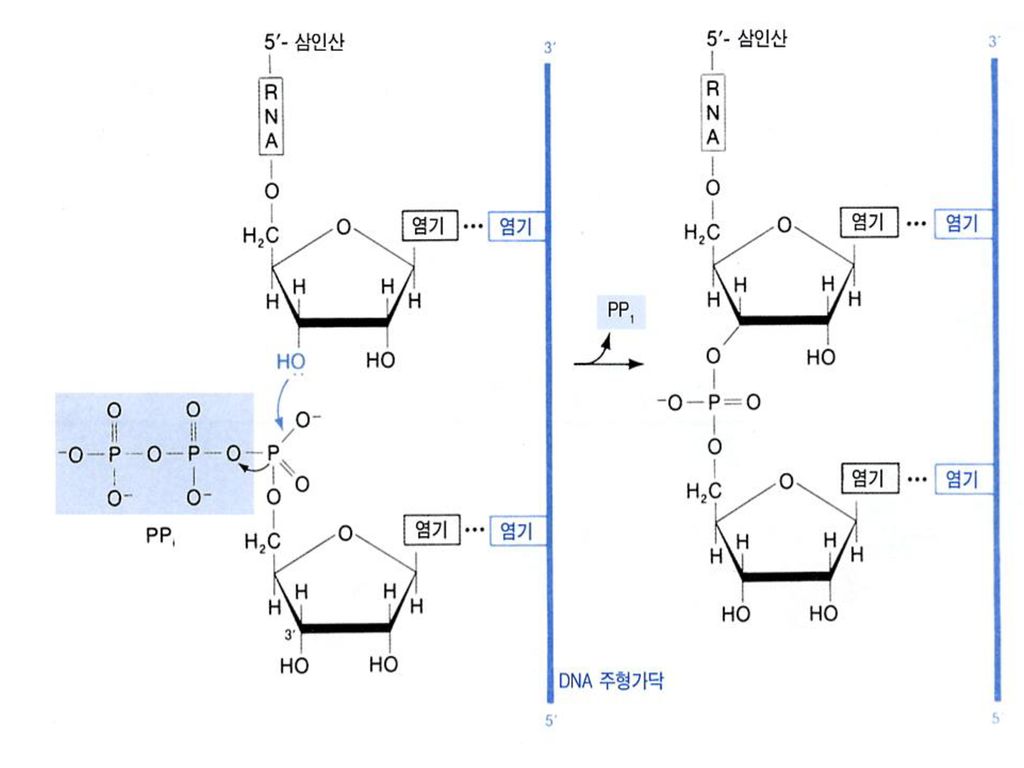
b. Elongation The release of the subunit causes
the conformational change of the core enzyme. The core enzyme slides on the DNA template toward the 3 end. Free NTPs are added sequentially to the 3-OH of the nascent RNA strand. (NMP)n + NTP (NMP)n+1 + PPi RNA strand substrate elongated RNA strand
n + NTP (NMP)n+1 + PPi. RNA strand substrate. elongated. RNA strand.")
52
RNA-pol, DNA segment of ~40nt and
the nascent RNA form a complex called the transcription bubble. The 3 segment of the nascent RNA hybridizes with the DNA template, and its 5end extends out the transcription bubble as the synthesis is processing.

55
R가 terminator에 결합

56
Transcription bubble

58
Protection from degradation
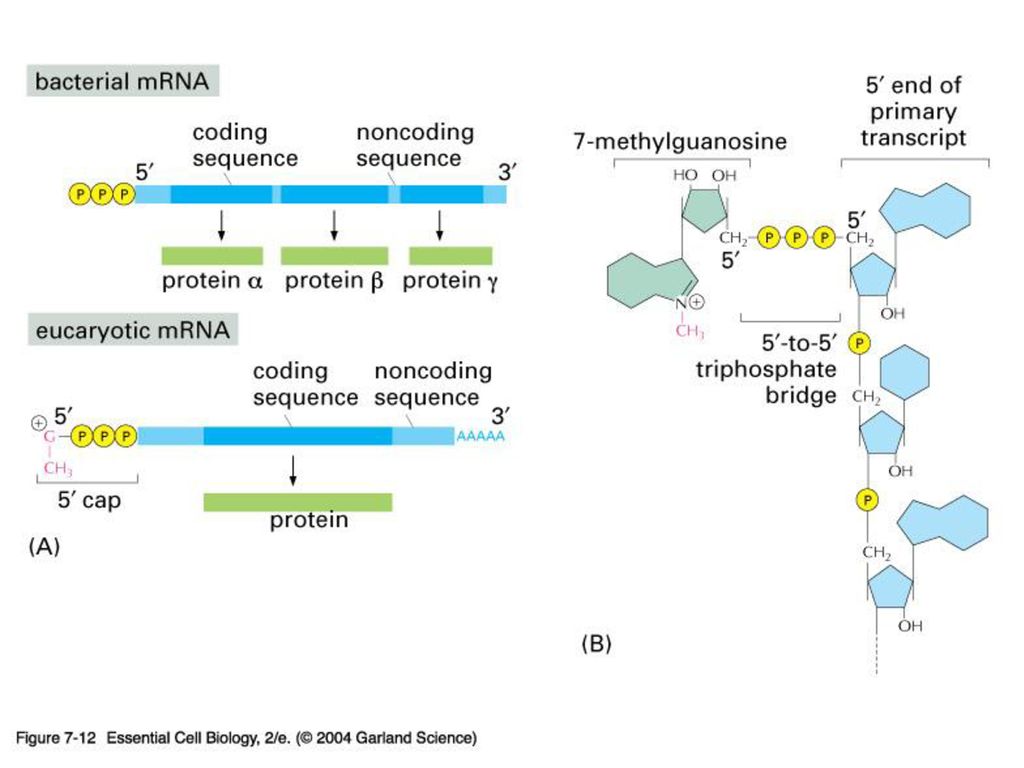
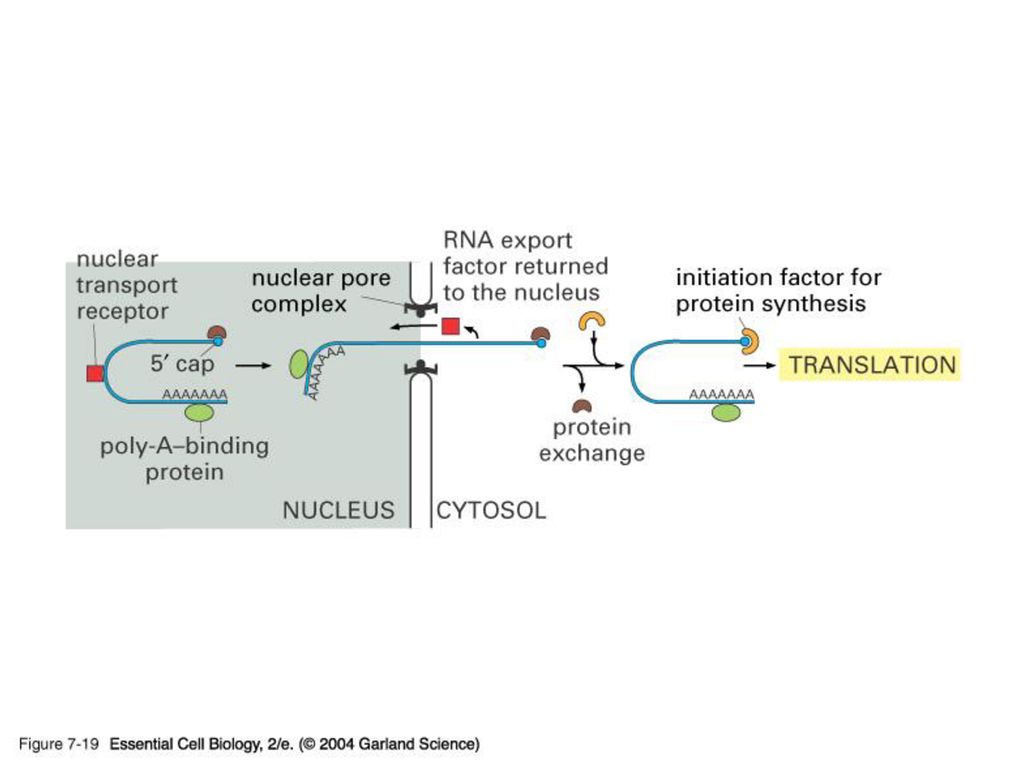
5’-capping과 poly(A) tail 의 기능 Protection from degradation (due to longer half life---think maternal transcripts) 방향 표지 Translation 조절에 관여 (고도로 분화된 진핵 생물 유전자 발현 조절의 한 단면)
 tail 의 기능. Protection from degradation. (due to longer half life---think maternal transcripts) 방향 표지. Translation 조절에 관여. (고도로 분화된 진핵 생물 유전자 발현 조절의 한 단면)")
60
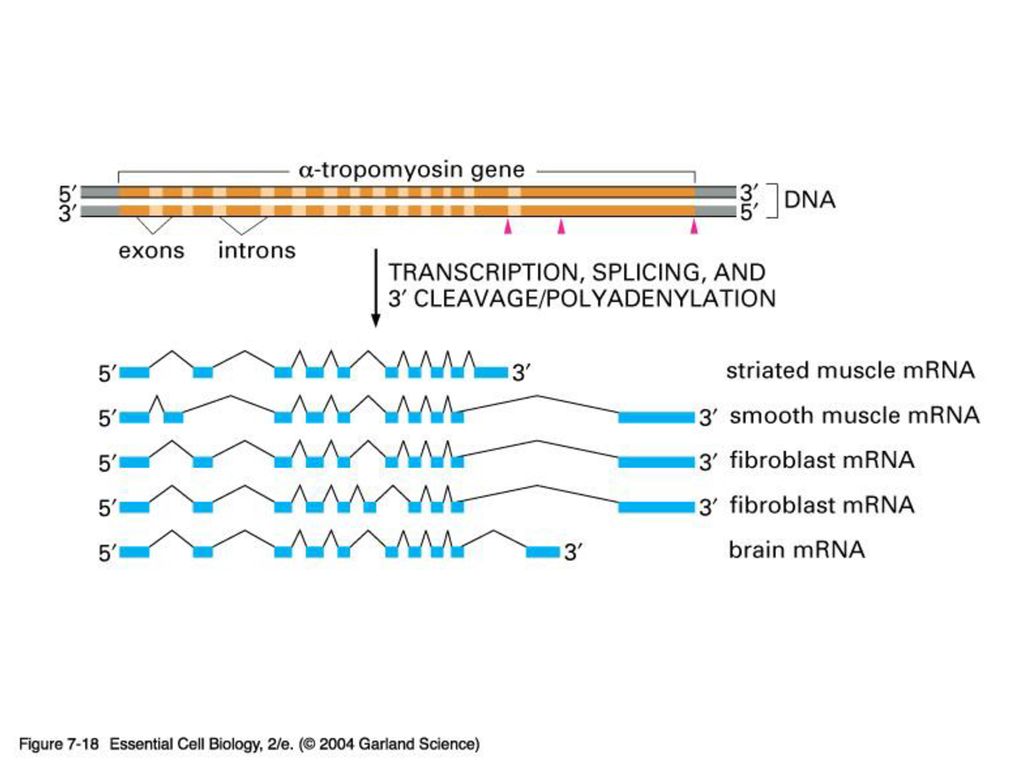
-제한된 숫자의 유전자를 다양하게 이용 할 수 있음 (differential splicing)
MIT의 Phil Sharp가 진핵생물의 intron을 발견한 공로로 1994년 Nobel 상 수상. Why intron?? -때로는 유전자 발현 조절에 관여 -제한된 숫자의 유전자를 다양하게 이용 할 수 있음 (differential splicing)
")
64
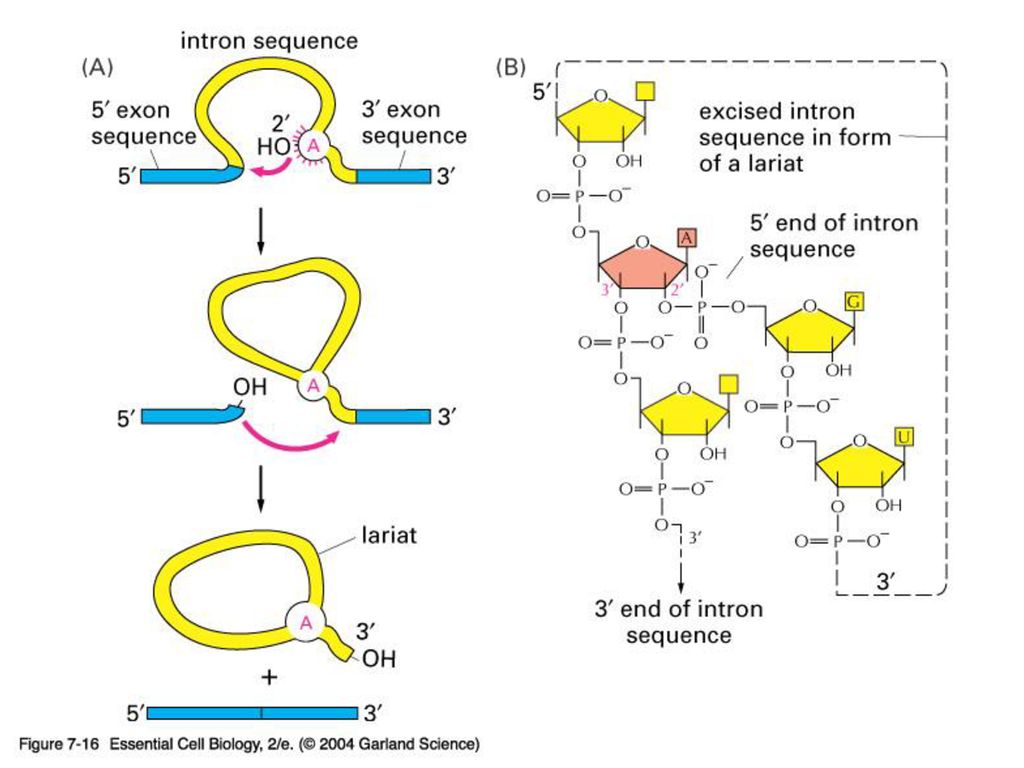
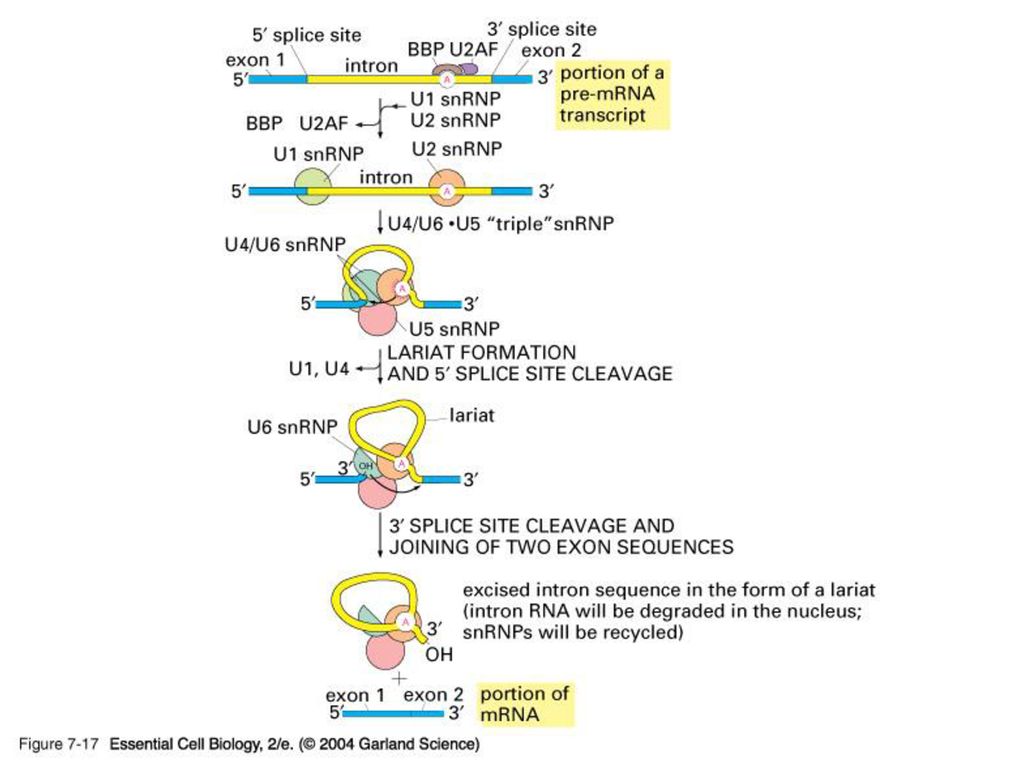
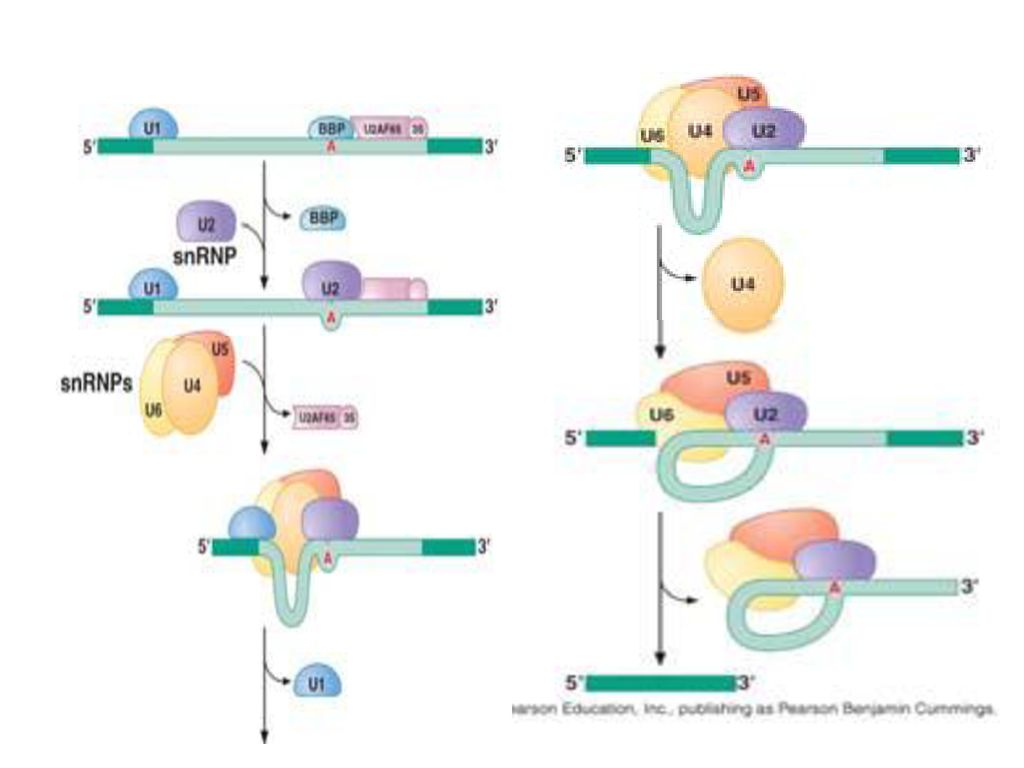
snRNPS (small nuclear ribonucleoprotein particles) ,
작은핵 RNA (snRNA)와 단백질의 복합체 “Splisosome”을 형성한다. RNA가 효소 작용을 하는 ribozyme의 대표적인 예. Other ribozymes: ribonuclease P ribosomal large subunit (28S rRNA) many more to come
와 단백질의 복합체. Splisosome 을 형성한다. RNA가 효소 작용을 하는 ribozyme의 대표적인 예. Other ribozymes: ribonuclease P. ribosomal large subunit (28S rRNA) many more to come.")
69
Splicing

70
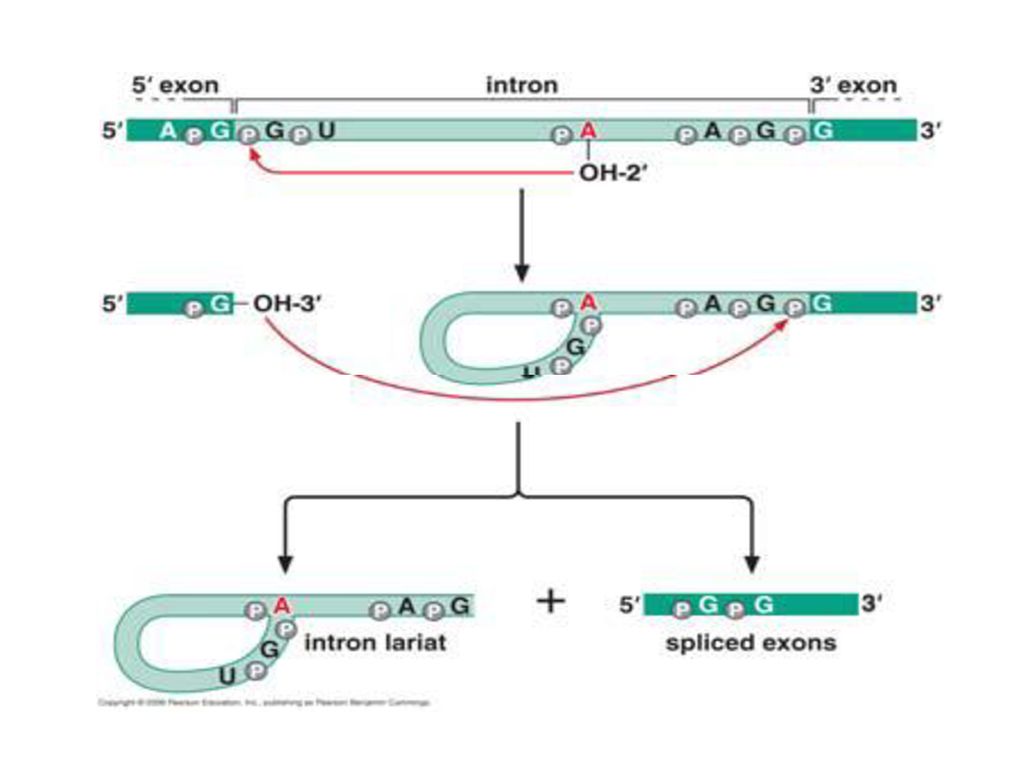
Exon and intron Exons are the coding sequences that appear on split genes and primary transcripts, and will be expressed to matured mRNA. Introns are the non-coding sequences that are transcripted into primary mRNAs, and will be cleaved out in the later splicing process.

71
Splicing mechanism

73
Trans-splicing

74
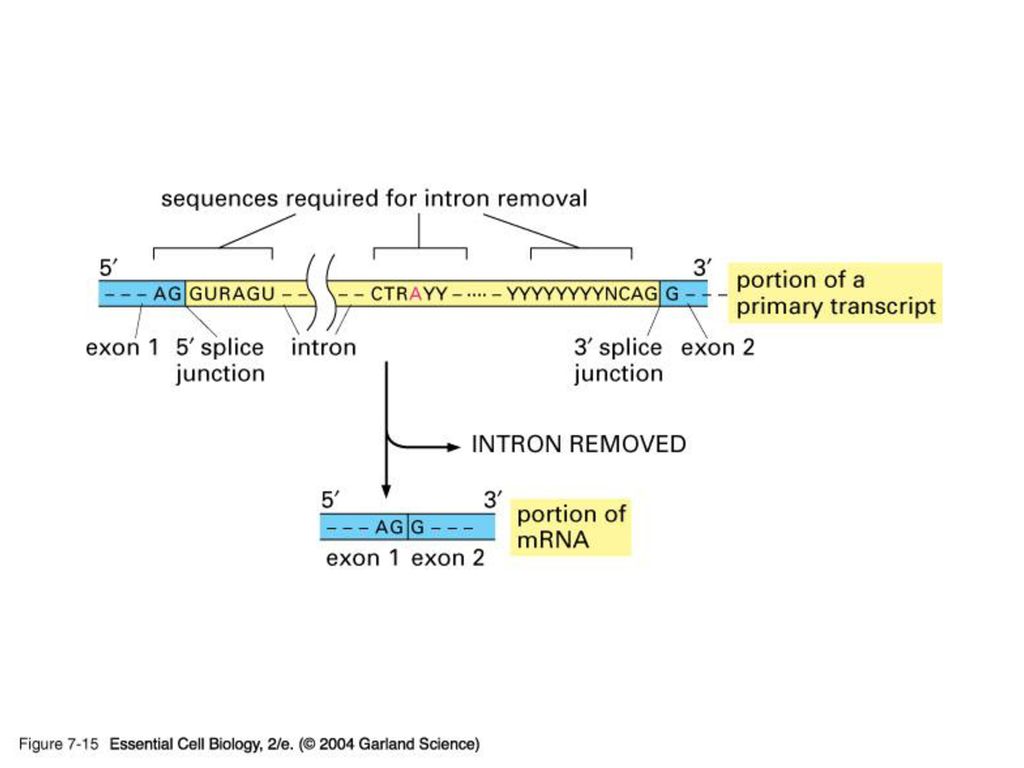
Spliceosome Splicing introns from mRNA occurs at short, conserved sequences called splice sites which specify the beginning and ends of introns. GU on the 5' end and AG on the 3' end are 100% conserved. Catalyzed by a structure called a spliceosome, composed of protein and RNA (snRNA – small nuclear RNA)
")
76
Three classes of RNA splicing

77
Converting Group I introns into ribozymes

78
RNA-RNA hybrids during splicing

79
Errors by mistakes in splice-site selection

80
The AT-AC spliceosome catalyzed splicing

81
Alternative splicing

82
전사 (transcription) DNA-dependent RNA polymerase의 작용으로 DNA를 주형으로 RNA 가닥을 합성하는 과정. rNTP를 building block으로 사용하며 연결 과정에서 PPi가 빠져 나오는 것, 그리고 새로운 사슬의 합성이 5’ 3’으로 진행되는 것은 DNA복제와 마찬가지. DNA 복제 ( replication) 과의 차이점: -두 가닥의 주형 DNA 중 한 가닥만 읽는다 (만들어진 산물도 외가닥) - 일부 지역만 읽는다 (만들어진 산물의 길이가 짧다) - Thymine이 올 자리에 대신 uracil이 들어간다. - 선도 사슬이 필요없이 끝자리에서부터 시작 할 수 있다. - 교정 작업 (proof reading) 을 하지 않는다.
 과의 차이점: -두 가닥의 주형 DNA 중 한 가닥만 읽는다. (만들어진 산물도 외가닥) - 일부 지역만 읽는다. (만들어진 산물의 길이가 짧다) - Thymine이 올 자리에 대신 uracil이 들어간다. - 선도 사슬이 필요없이 끝자리에서부터 시작 할 수 있다. - 교정 작업 (proof reading) 을 하지 않는다.")
83
왜 우리는 실제 주형과 상보적인 가닥을 읽을까?

84
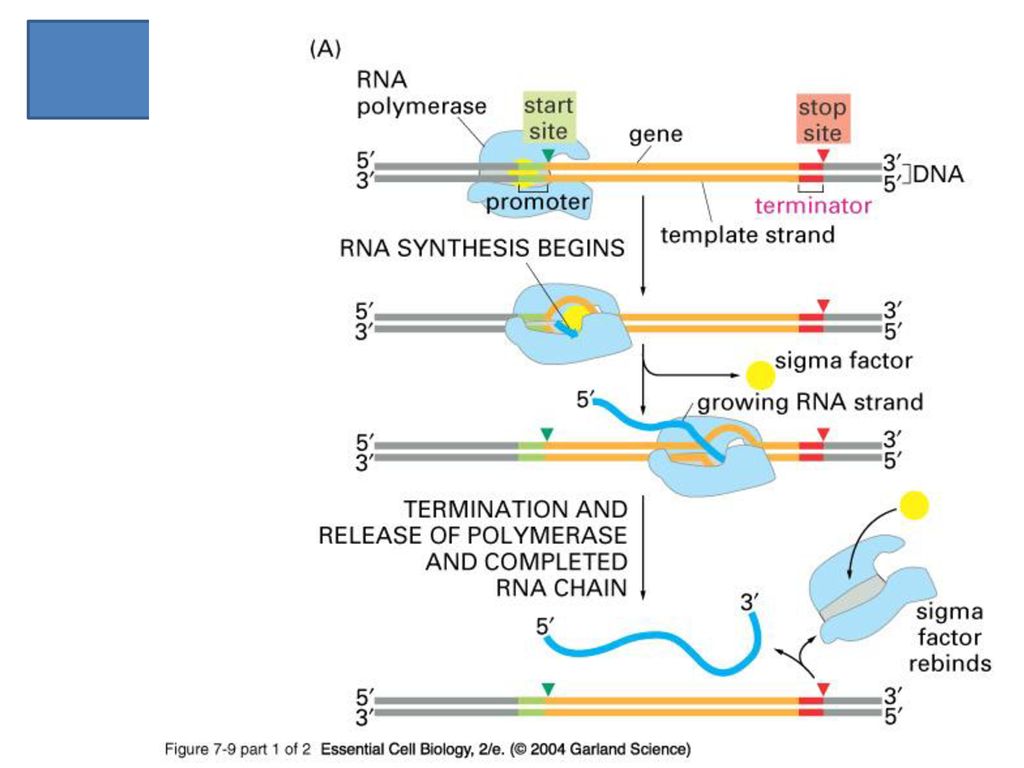
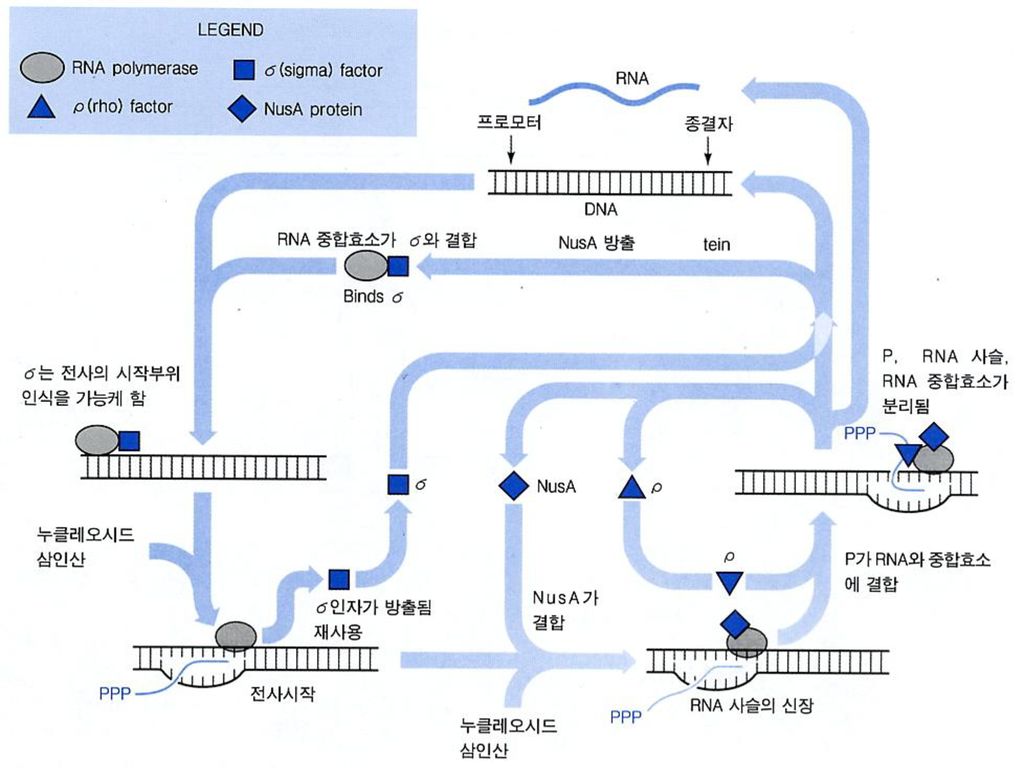
Transcription, 그 중에서도 mRNA의 합성 과정에 관한 분자생물학적 지식들은 먼저 간단한 대장균을 통하여 밝혀짐.
DNA에 부착하여 RNA 를 합성하고 있는 대장균의 RNA 중합효소 ( 2 a + b + b’ ) + s -core enzyme (핵심효소) -holoenzyme (전효소) 유전자가 앞쪽에 s-factor 가 달라붙는 부위가 있어 그 바로 뒷부분에서부터 전사가 시작된다..
 + s. -core enzyme (핵심효소) -holoenzyme (전효소) 유전자가 앞쪽에 s-factor 가 달라붙는 부위가 있어 그 바로 뒷부분에서부터 전사가 시작된다..")
85
Alias “TATA” box 진핵생물에서는 위치가 일정치 않음

86
TTGACA TATAAT

87
RNA 합성의 4 단계 개방 복합체의 형성 개시 (initiation) 사슬의 연장 (elongation)
종결 (termination)
")
88
Open-promoter complex
DNA 복제의 경우 helicase가 담당하는 작업. RNA 전사때에는 RNA polymerase가 primerase + helicase 의 역할도 담당. 전사가 시작될 부위를 인지하는 것은 s-factor. Initiation 항상 그런 것은 아니지만 주로 A 또는 G 에서 시작된다. Elongation RNA 중합효소 복합체가 한 자리씩 옮겨 가면서 새로 들어오는 rNTP를 이제 까지 만들어 진 사슬에 붙여 나간다. ( next slide)
")
93
진핵 세포의 RNA는 합성 후 핵에서 공정 단계를 거치고 난 뒤 세포질로
방출된다. (5’-capping, 2nd 염기의 methylation, polyadenylation, splicing)
")
94
5’-capping과 poly(A) tail 의 기능
Protection from degradation (due to longer half life---think maternal transcripts) 방향 표지 Translation 조절에 관여 (고도로 분화된 진핵 생물 유전자 발현 조절의 한 단면)
 방향 표지. Translation 조절에 관여. (고도로 분화된 진핵 생물 유전자 발현 조절의 한 단면)")
96
INTRON

97
MIT의 Phil Sharp가 진핵생물의 intron을 발견한 공로로
1994년 Nobel 상 수상. Why intron?? -때로는 유전자 발현 조절에 관여 -제한된 숫자의 유전자를 다양하게 이용 할 수 있음 (differential splicing)
")
98
snRNPS (small nuclear ribonucleoprotein particles) ,
작은핵 RNA (snRNA)와 단백질의 복합체 “Splisosome”을 형성한다. RNA가 효소 작용을 하는 ribozyme의 대표적인 예. Other ribozymes: ribonuclease P ribosomal large subunit (28S rRNA) many more to come
와 단백질의 복합체. Splisosome 을 형성한다. RNA가 효소 작용을 하는 ribozyme의 대표적인 예. Other ribozymes: ribonuclease P. ribosomal large subunit (28S rRNA) many more to come.")
99
Translation

100
단백질 합성의 세 기구: 1) mRNA-the template 2) tRNA-the delieverer 3) rRNA (ribosome)-the synthesizer
 mRNA-the template 2) tRNA-the delieverer 3) rRNA (ribosome)-the synthesizer")
101
1950년대 말 까지 DNA가 일단 RNA로 전사 된 뒤 이로부터 단백질이 합성된다는 것에는 의견이 모아짐.
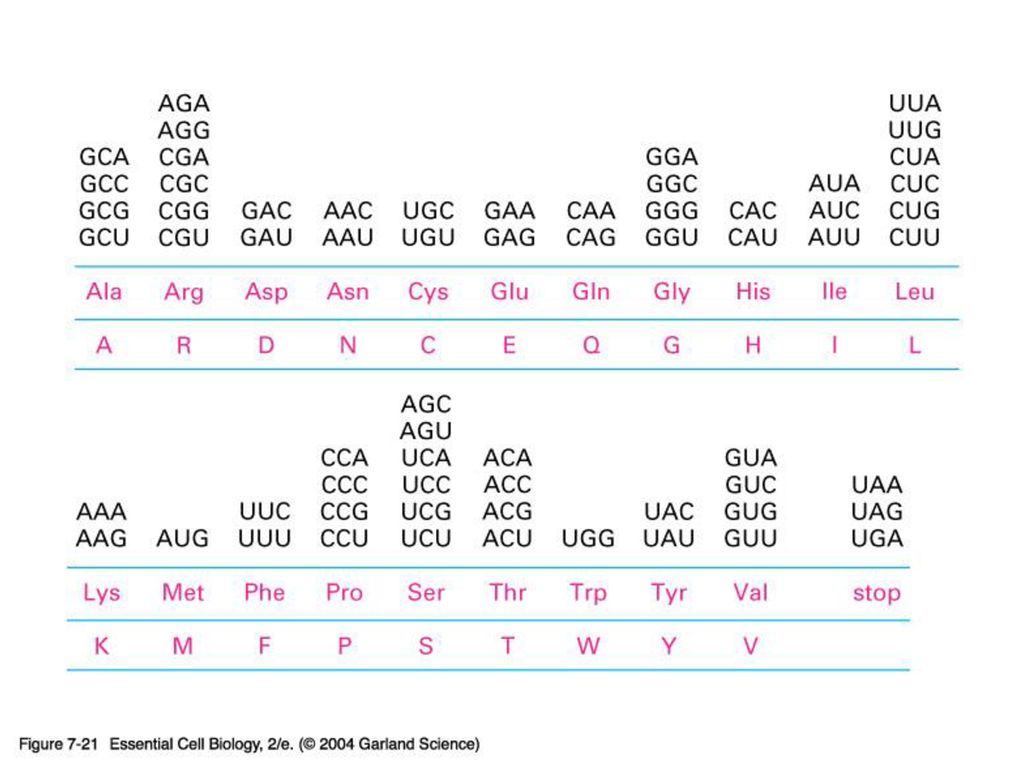
1) mRNA 1950년대 말 까지 DNA가 일단 RNA로 전사 된 뒤 이로부터 단백질이 합성된다는 것에는 의견이 모아짐. The question is HOW? RNA(=핵산)의 언어가 단백질의 언어로 바뀌는 과정은 일종의 암호해독(deciphering)을 필요로 한다. Code – coding – triplet codon 하나의 아미노산을 stand for 하는 핵산이 세 개 (triplet codon)이라는 것을 어떻게 알아내었나?? 핵산의 종류는 4개. 아미노산의 종류는 20개. if mono-codons: 가능한 아미노산의 종류는 only 4 개. if doublet –codons : 가능한 아미노산의 종류는 4 X 4 = only 16 개 따라서 최소한 3개의 핵산이 하나의 아미노산을 암호화 해야 함.
 mRNA. 1950년대 말 까지 DNA가 일단 RNA로 전사 된 뒤 이로부터 단백질이 합성된다는 것에는 의견이 모아짐. The question is HOW RNA(=핵산)의 언어가 단백질의 언어로 바뀌는 과정은 일종의 암호해독(deciphering)을 필요로 한다. Code – coding – triplet codon. 하나의 아미노산을 stand for 하는 핵산이 세 개 (triplet codon)이라는 것을 어떻게 알아내었나 핵산의 종류는 4개. 아미노산의 종류는 20개. if mono-codons: 가능한 아미노산의 종류는 only 4 개. if doublet –codons : 가능한 아미노산의 종류는 4 X 4 = only 16 개. 따라서 최소한 3개의 핵산이 하나의 아미노산을 암호화 해야 함.")
106
2) tRNA
 tRNA")
107
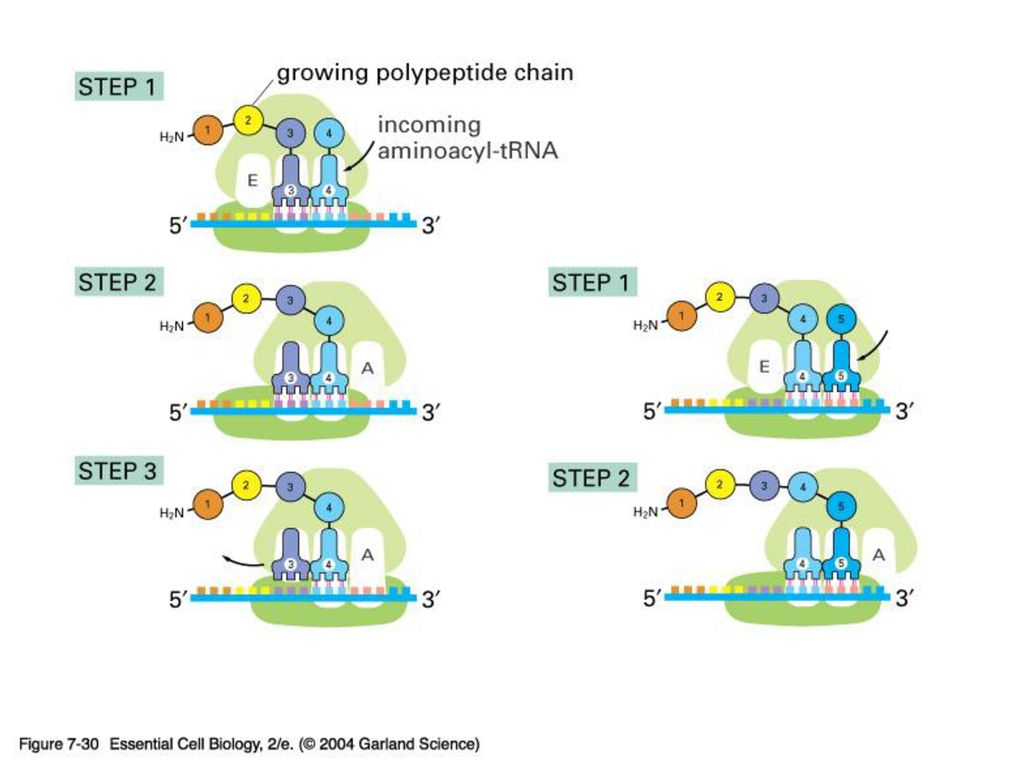
tRNA는 mRNA를 해독하면서 해당 아미노산과 연결해주는 adapter 이다.
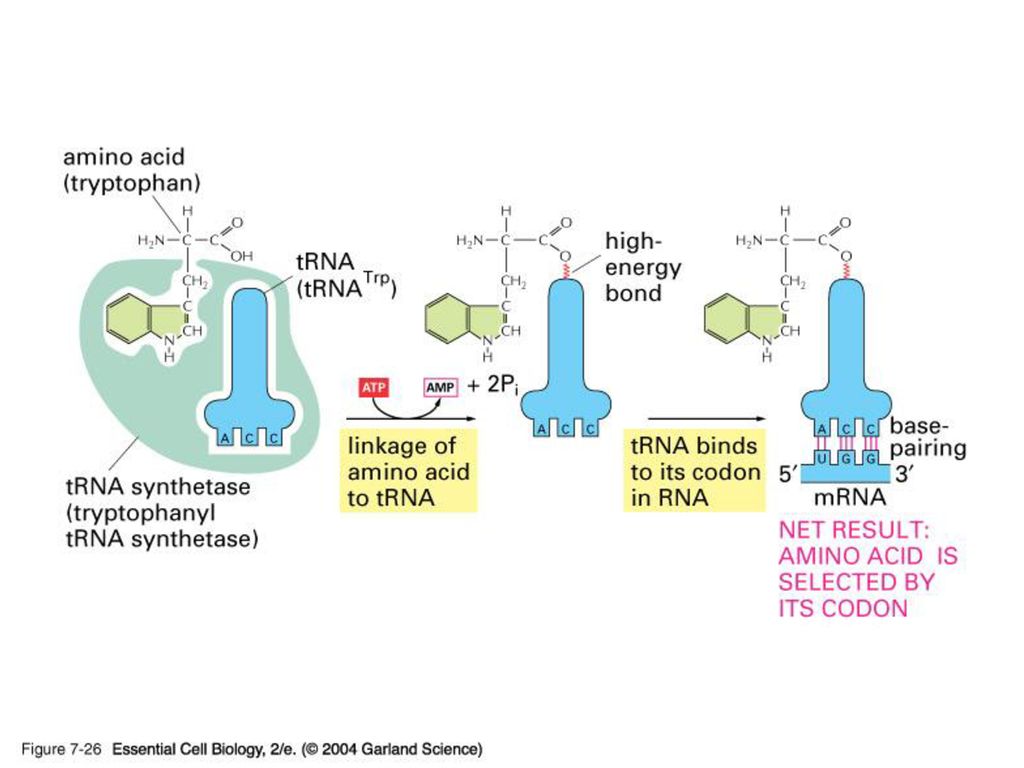
약 80 nt로 이루어진 작은 RNA 가닥. 2차구조로 clover leaf모양을 형성하며 두 개의 leaf가 겹쳐서 다시 L자 모양. 두가닥을 이루지 않는 두 부위중 하나는 RNA상의 triplet codon과 결합하는 anticodon, 그리고 다른 하나는 해당 아미노산이 부착되어 있는 자리이다. 각각의 tRNA에 누가 지정된 amino acid를 붙여주나? 각 amino acid 마다 정해진 aminoacyl – tRNA – synthetase가 있다. 모두 20개. Anticodon과 아미노산 수용기 팔의 염기서열을 보고 판독. 따라서 translation 과정에서 이 효소의 정확도는 매우 중요. (A number game) 20 aminoacyl-tRNA synthetase 31 tRNA 61 (64-3 stop) codon 이것은 한 synthetase가 하나 이상의 tRNA를, 그리고 한 tRNA가 여러 개의 codon을 인지 할 수 있음을 보여준다.
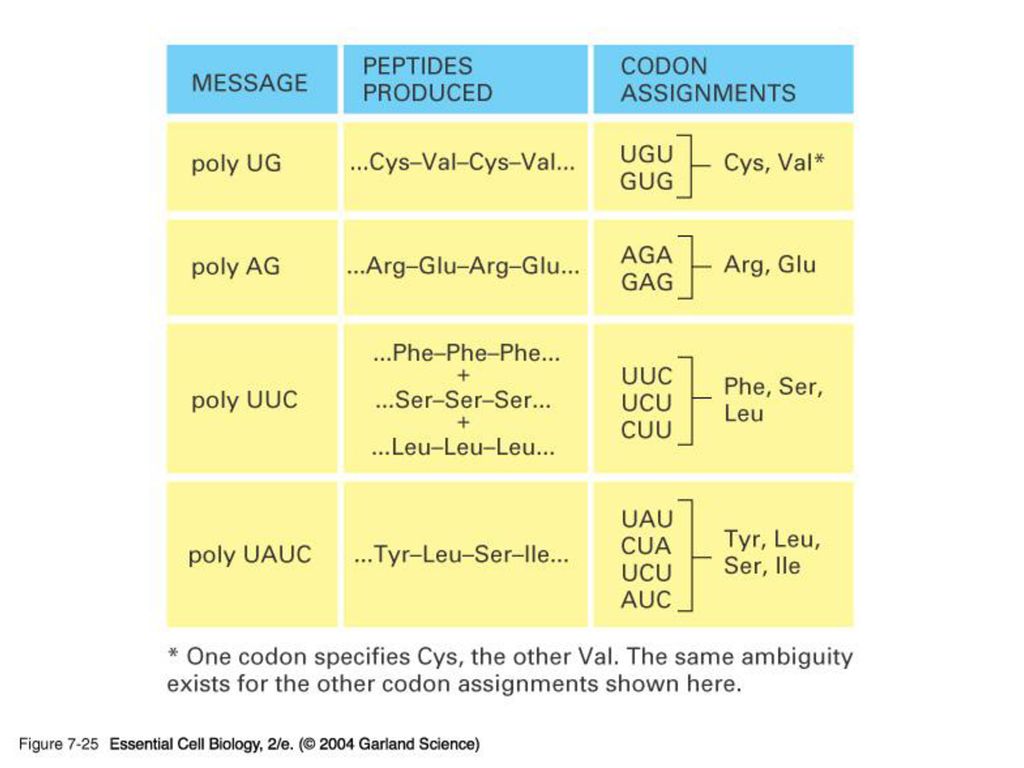
 20 aminoacyl-tRNA synthetase. 31 tRNA. 61 (64-3 stop) codon. 이것은 한 synthetase가 하나 이상의 tRNA를, 그리고 한 tRNA가 여러 개의 codon을 인지 할 수 있음을 보여준다.")
109
tRNA의 구조상 anticodon의 첫번째 (=codon의 세번째) nucleotide는 완벽한 수소결합이 형성되지 않아도 좋다.
따라서 codon의 세 번째 자리에 약간의 변이가 허용됨. (=wobble) 바로 이 현상 때문에 많은 자연발생적 점돌연변이가 치명적인 결과를 가져오지 않고 넘어갈 수 있다. 또한 한 aminoacid에 배당되는 염기서열이 두 개 이상인 것도 이 현상으로 설명된다. (p. 241 그림 21 다시보기)
 바로 이 현상 때문에 많은 자연발생적 점돌연변이가 치명적인 결과를 가져오지 않고 넘어갈 수 있다. 또한 한 aminoacid에 배당되는 염기서열이 두 개 이상인 것도 이 현상으로 설명된다. (p. 241 그림 21 다시보기)")
110
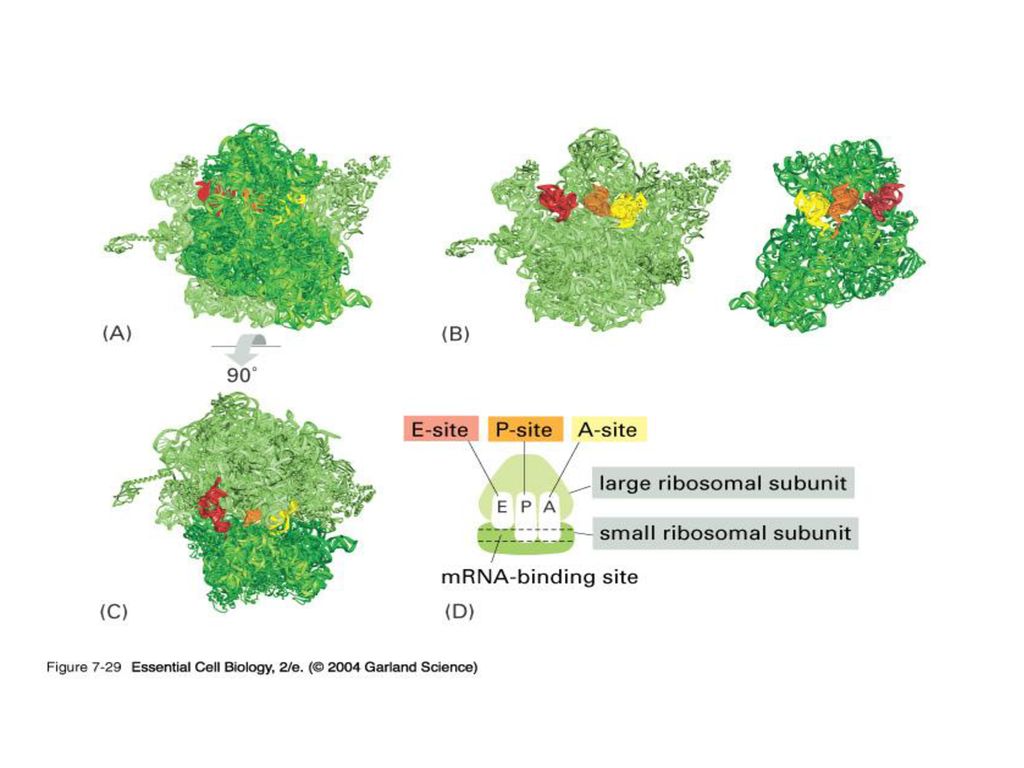
3) rRNA (ribosome) 실제로 단백질의 조립이 일어나는 공장.
Some ribosomes are free. Some are bound to the ER.

111
진핵생물의 ribosome은 세포질에서 합성된 rProtein이 핵으로 들어가 rRNA와 조합된 뒤 다시 세포질로 나와서 활동.
원핵생물은 large, small subunit가 모두 조금씩 작다.

112
원핵생물의 ribosome large subunit를 형성하는 rRNA (23S + 5 S
원핵생물의 ribosome large subunit를 형성하는 rRNA (23S + 5 S. 진핵생물의 경우는 세 개의 28S rRNA.) 그림에서 보듯이 ribosome의 large subunit는 RNA (not protein) 이 전체적인 모양을 결정하며 peptide bond를 형성하는 기능도 담당한다. A Ribozyme.
 그림에서 보듯이 ribosome의 large subunit는 RNA (not protein) 이 전체적인 모양을 결정하며 peptide bond를 형성하는 기능도 담당한다. A Ribozyme.")
116
3. Termination

117
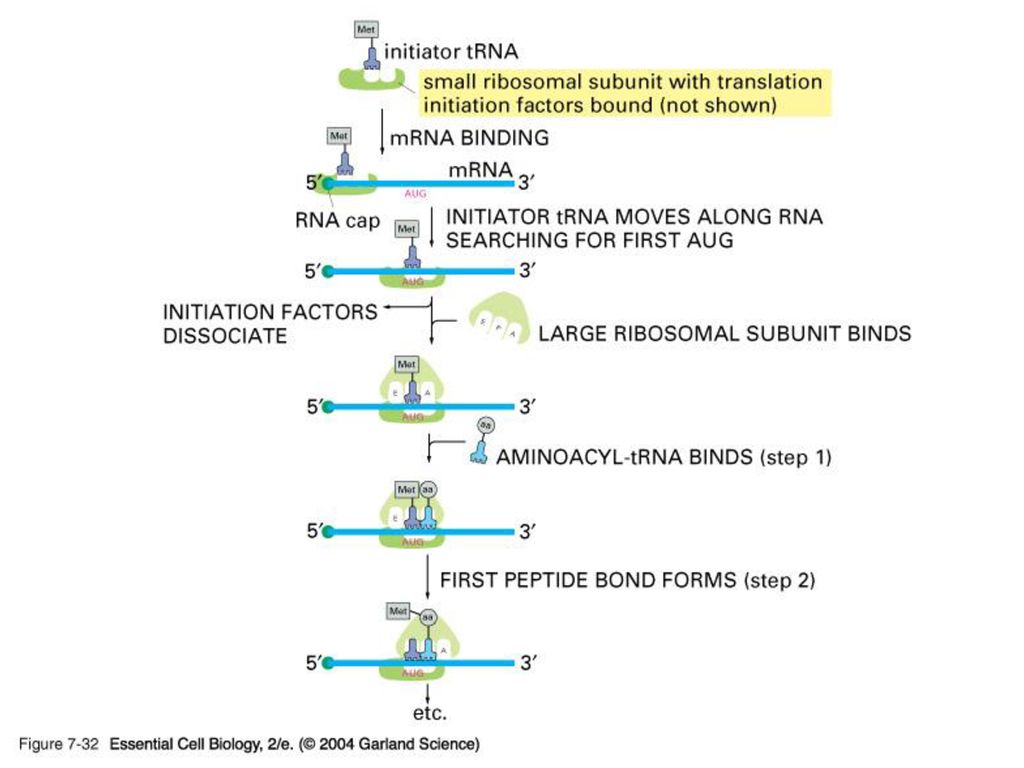
Shine-Dalgarno (SD) sequence : AGGAGGU
5’-cap이 없는 원핵생물은 어떻게 단백질 합성을 시작하나? 각 ORF 의 앞쪽에 위치한 SD를 16S rRNA가 인지하고 부착한다. 원핵생물의 rRNA small subunit도 initiator tRNA를 장착하고 다님. 단 이 경우는 보통 Met가 아닌 fMET. 왜일까??

118
Polyribosome 또는 polysome
진핵과 원핵생물에 모두 존재. 단 원핵생물에서는 mRNA가 아직 DNA에 붙어있는 상태에서 polysome이 형성된다.

119
진핵생물과 원핵생물의 단백질 합성 과정에서 찾을 수 있는 세부적인 차이는 인간에게 편리하다.
e.g. 원핵생물의 단백질 합성만을 저해하는 인자를 사용하면 우리 몸의 세포에는 해를 주지 않으면서 감염된 병원균만을 죽일 수 있음. 이것이 바로 항균제, 또는 항생제 (antibiotics). 알려진 대부분의 항생제는 원핵생물의 단백질 합성단계중 하나를 저해한다. (p. 252 표 7-2) 곰팡이나 균류, 또는 원생동물 (공통점: 진핵생물!)이 일으키는 질병을 치료하기 어렵고 또한 이들을 죽이기 위하여 사용되는 약물이 환자에게 고통을 주는 이유는??
. 알려진 대부분의 항생제는 원핵생물의 단백질 합성단계중 하나를 저해한다. (p. 252 표 7-2) 곰팡이나 균류, 또는 원생동물 (공통점: 진핵생물!)이 일으키는 질병을 치료하기 어렵고 또한 이들을 죽이기 위하여 사용되는 약물이 환자에게 고통을 주는 이유는")
120
합성된 단백질의 분해 세포내에서 만들어진 단백질을 신속히 분해해야 하는 경우들이 있다.
-특정 상황에서만 필요한 단백질 (세포주기 조절인자, 신호전달물질) 잘못 접힌 단백질 이를 위하여 전문적인 분해 장소 (=proteasome) 가 존재. Ubiquitin 에 의한 단백질 분해 (ubiquitin-dependent proteolysis)는 가장 대표적인 단백질 분해 기구 .
 -잘못 접힌 단백질 이를 위하여 전문적인 분해 장소 (=proteasome) 가 존재. Ubiquitin 에 의한 단백질 분해 (ubiquitin-dependent proteolysis)는 가장 대표적인 단백질 분해 기구 .")
121
세포내에서 만들어진 단백질을 신속히 분해해야 하는 경우들이 있다.
합성된 단백질의 분해 세포내에서 만들어진 단백질을 신속히 분해해야 하는 경우들이 있다. -특정 상황에서만 필요한 단백질 (세포주기 조절인자, 신호전달물질, etc) -잘못 접힌 단백질 이를 위하여 전문적인 분해 장소 (=proteasome) 가 존재. Ubiquitin 에 의한 단백질 분해 (ubiquitin-dependent proteolysis)는 가장 대표적인 단백질 분해 기구 .
 -잘못 접힌 단백질. 이를 위하여 전문적인 분해 장소 (=proteasome) 가 존재. Ubiquitin 에 의한 단백질 분해 (ubiquitin-dependent proteolysis)는 가장 대표적인 단백질 분해 기구 .")
123
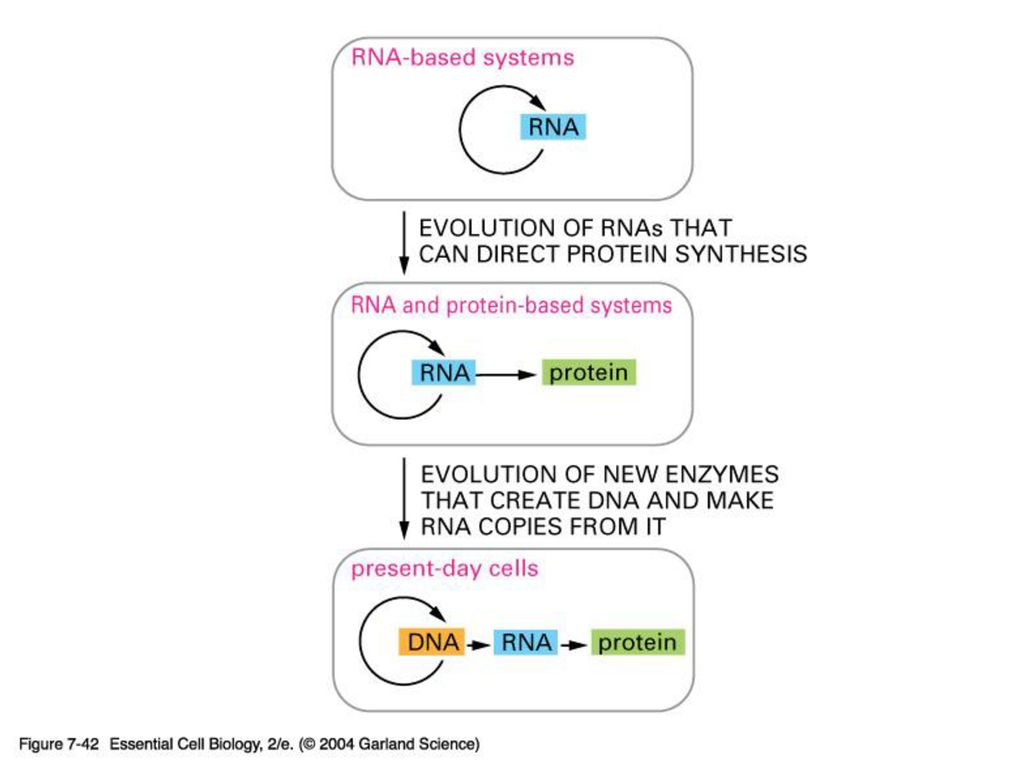
The RNA-World Hypothesis

124
Idea의 저장 Idea의 실행 매개자

125
1. RNA가 DNA보다 먼저 만들어졌다? 2. 실제 RNA가 유전물질로 작용할 수 있다
RNA world 가설을 지지하는 실제 증거들 1. RNA가 DNA보다 먼저 만들어졌다? -핵산의 모든 염기 (아데닌, 구아닌 시토신)은 각기 고유의 생합성 과정을 거치는 반면 유독 티민만은 우라실의 methylation을 통하여 만들어 진다. - 프라이머가 있어야만 하는 DNA 중합효소와는 달리 RNA중합효소는 새로운 가닥을 시작할 수 있다. 2. 실제 RNA가 유전물질로 작용할 수 있다 현재도 일부 바이러스는 RNA를 유전 물질로 사용한다. RNA virus중 retrovirus는 어쩌면 유전정보의 저장 기능이 RNA에서 DNA로 이양되는 과정을 보여주는 중간단계가 아닐런지?
은 각기 고유의 생합성 과정을 거치는 반면 유독 티민만은 우라실의 methylation을 통하여 만들어 진다. - 프라이머가 있어야만 하는 DNA 중합효소와는 달리 RNA중합효소는 새로운 가닥을 시작할 수 있다. 2. 실제 RNA가 유전물질로 작용할 수 있다. 현재도 일부 바이러스는 RNA를 유전 물질로 사용한다. RNA virus중 retrovirus는 어쩌면 유전정보의 저장 기능이 RNA에서 DNA로 이양되는 과정을 보여주는 중간단계가 아닐런지")
126
RNA가 단백질의 역할을 맡는다는 증거 - 핵 속에서 여러 가지 작은 non-coding RNA들이 intron의 제거, mRNA의 변형과 교정, 그리고 telomerase에 의한 말단 연장에 사용된다.) - Ribosome 내에서 아미노산과 아미노산 사이의 peptide결합은 rProtein이 아니라 rRNA에 의하여 촉매된다. - RibonucleaseP는 tRNA의 전구체를 잘라서 완성된 형태로 만들어준다.

128
2) RNA can modify other RNA
 RNA can modify other RNA")
129
3) RNA can catalyze its own synthesis
 RNA can catalyze its own synthesis")
131
여러 가지 non-coding RNA RNA 이름 기능 rRNA ribosomal RNA 단백질 합성 tRNA transfer RNA 아미노산 운반 snRNA small nuclear RNA splicing snoRNA small nucleolus(인) RNA RNA processing aRNA antisense RNA 유전자 발현 조절 miRNA micro RNA 유전자 발현 조절 siRNA small interfering RNA 유전자 발현 조절
 RNA RNA processing. aRNA antisense RNA 유전자 발현 조절. miRNA micro RNA 유전자 발현 조절. siRNA small interfering RNA 유전자 발현 조절.")
132
유전자에서 단백질까지 요약1

133
유전자에서 단백질까지 요약2


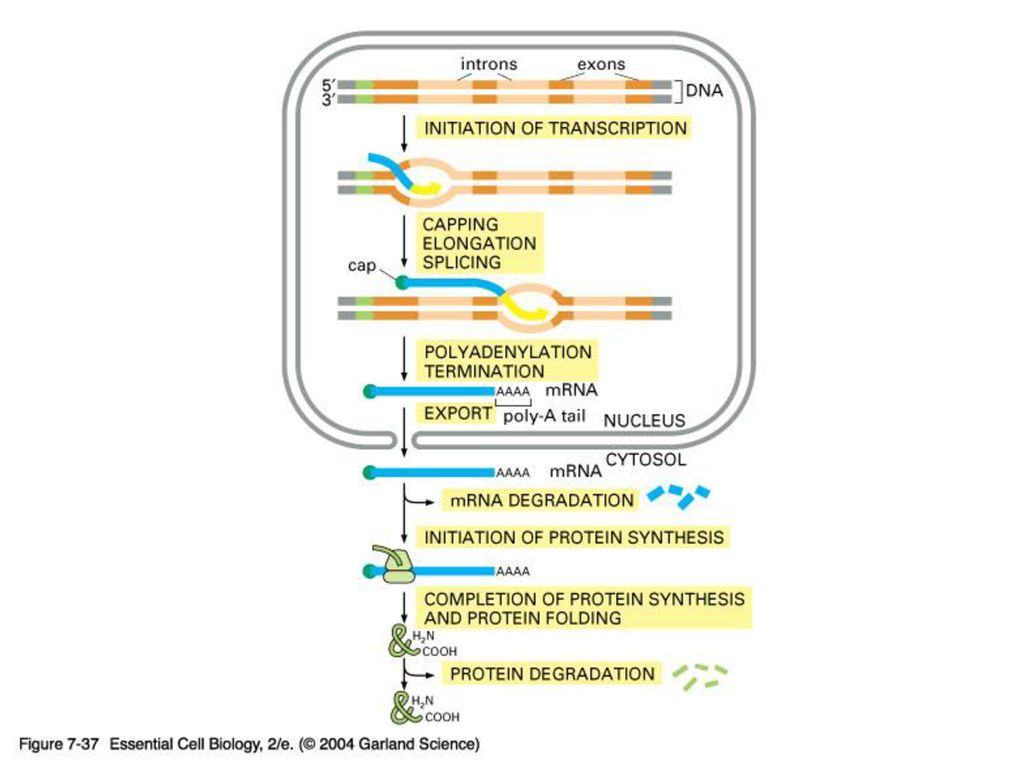




. Polymerase Chain Reaction (PCR) 중합효소연쇄반응 주형 DNA 분자의 특정 부분을 선택적으로 증폭 Primer set 와 내열성 DNA polymerase 를 이용하여 짧은 시간 내에 미량의 시료에서 Kary.>")

>")
 RNA 합성 가공 발현 조절.>")











>")