Download presentation
3
DNA & Double Helix

4
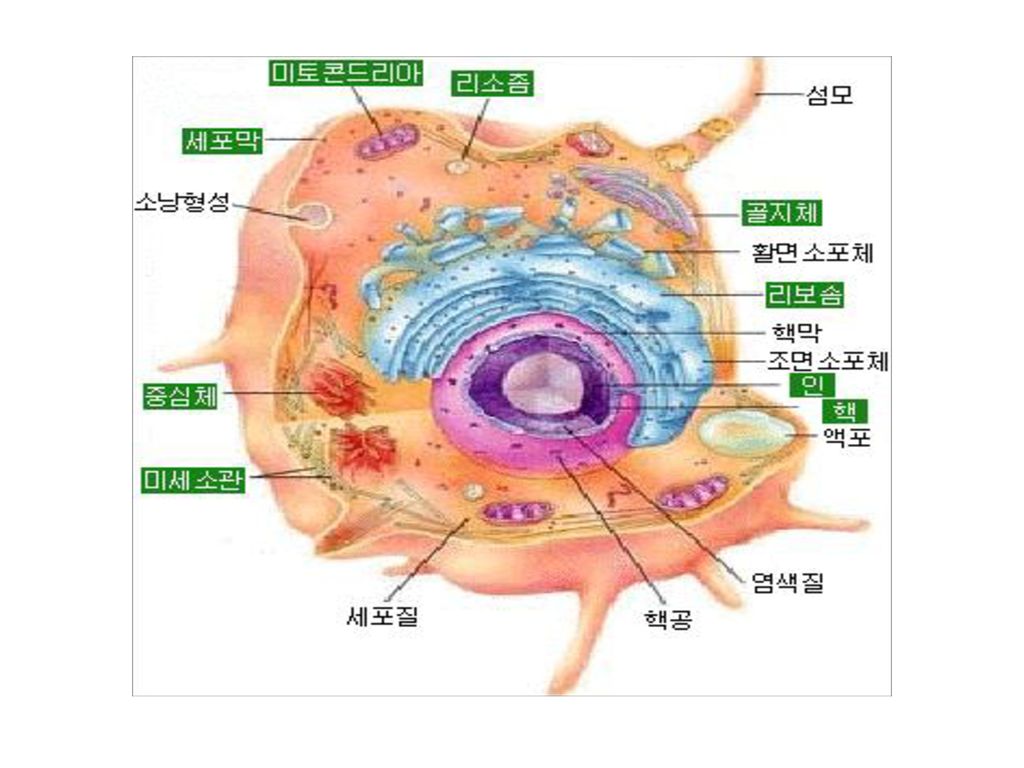
3 유전학의 놀라운 비밀 - DNA! 진핵세포에 있어서의 유전정보의 흐름

5
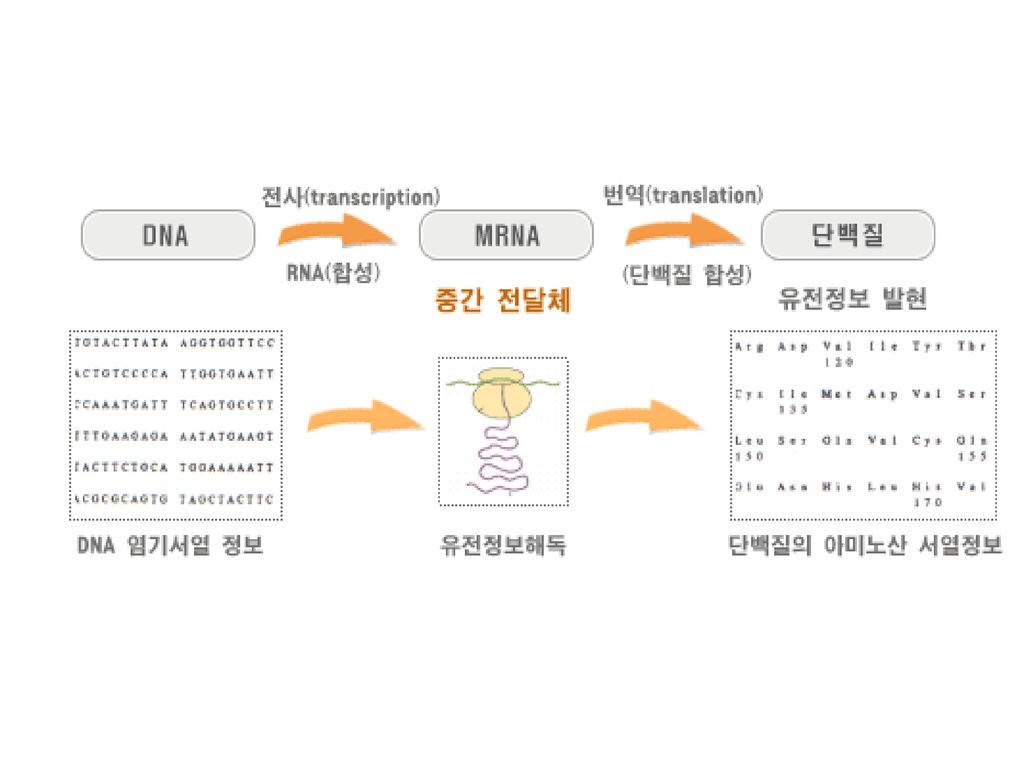
3 유전학의 놀라운 비밀 - DNA! 전사- DNA - > RNA

7
DNA 구조 연구의 선구자들 Erwin Chagarff Linus Pauling Rosalind Franklin
Mauris Wilkins James Dewey Watson Francis Crick

8
DNA의 발견 미셔는 환자의 고름이 묻은 천에서 Nuclein(DNA 포함 물질)을 발견하였다.
을 발견하였다.")
9
Erwin Chagarff A-T, G-C의 염기만이 서로 결합한다는 상보성을 보여주는 실험 결과 발표

10
Linus Pauling DNA는 나선의 모양임을 발견

11
Franklin & Wilkins X선 회절법으로 DNA의 규격을 밝힘

12
Watson & Crick DNA가 이중나선임을 밝힘 DNA 구조 모형 제작

13
왓슨과 크릭은 1953년 ‘Molecular Structure of Nucleic Acids’란 제목의 논문에서 DNA 구조를 밝힘으로써 노벨상을 받았다.

15
DNA의 구조 DNA와 Nucleotide 염기(Base) 상보성 DNA 구조 다른 형태의 DNA
 상보성 DNA 구조 다른 형태의 DNA")
16
DNA란 무엇인가? DNA(Deoxyrobonucleic Acid) 모든 생물에 존재하며 유전현상을 담당하는 물질
사슬 모양의 정보 고분자 물질 여러 개의 뉴클레오티드(Nucleotide)로 구성 원소:탄소, 산소, 질소, 인, 수소
로 구성. 원소:탄소, 산소, 질소, 인, 수소.")
17
뉴클레오티드(Nucleotide) 염기(Base), 당(Sugar), 인산(Phosphate)으로 구성
나선의 1회전은 10개의 뉴클레오티드로 형성 Base+Backbone

18
뉴클레오티드의 종류

19
염기(Bases) 질소를 포함 염기성 퓨린(Purine) 6각형고리+5각형고리 피리미딘(pyrimidine) 하나의 6각형고리
 질소를 포함 염기성 퓨린(Purine) 6각형고리+5각형고리 피리미딘(pyrimidine) 하나의 6각형고리")
20
퓨린(Purine) 아데닌(Adenine) 구아닌(Guanine)
 아데닌(Adenine) 구아닌(Guanine)")
21
피리미딘(pyrimidine) 시토신(Cytosine) 티민(Thymine) 또는 우라실(Uracil)
 시토신(Cytosine) 티민(Thymine) 또는 우라실(Uracil)")
22
DNA의 구조 핵산 : DNA(2중나선), RNA( 단일사슬) 1. 구 조 : 염기-당-인산 : 리보오스(RNA) : 데옥시리보오스 DNA deoxyribose (산소가 없다는 뜻) DNA의 구조

23
DNA와 RNA

24
DNA의 구조

25
샤가프의 실험

26
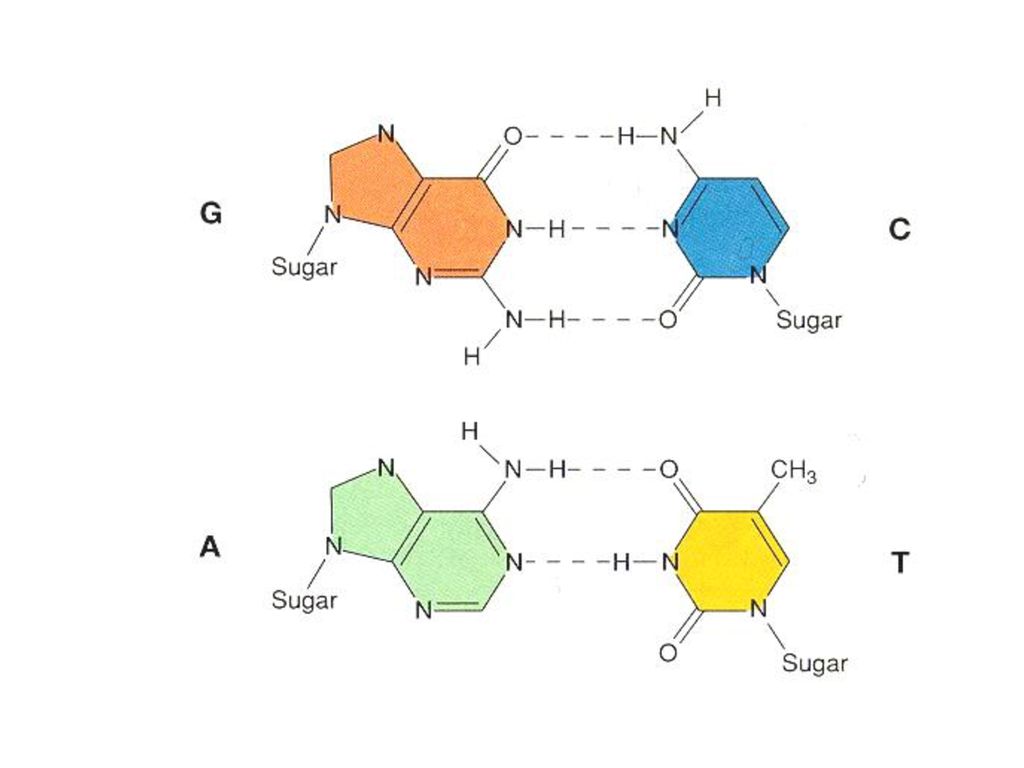
샤가프의 법칙 같은 생물종이 지닌 퓨린의 양(A+G)과 피리미딘의 양(T+C)은 같다.
A와 T의 양이 같고, G와 C의 양이 거의 같다. 즉 아데닌(A) : 티민(T) = 1 : 1이고, 구아닌(G) : 시토신(C) = 1 : 1인 것이다.
 : 티민(T) = 1 : 1이고, 구아닌(G) : 시토신(C) = 1 : 1인 것이다.")
27
상보결합의 규칙 한쪽 사슬의 염기배열을 결정하면, 다른쪽 사슬의 염기배열이 자동적으로 결정 A=T (아데닌-티민)
G≡C (구아닌-시토신)
")
28
A(아데닌)=T(티민)
=T(티민)")
29
DNA 구조의 특성 방향성 역평행(Antiparallel) 상보결합
 상보결합")
30
G(구아닌)≡C(시토신)
≡C(시토신)")
31
방향성 5’3’ 방향으로 결합 진행

32
역평행(Antiparallel) DNA의 두가닥 사슬은 서로 반대방향으로 되어있다.
 DNA의 두가닥 사슬은 서로 반대방향으로 되어있다.")
33
이중나선(Double Helix) 여러 개의 뉴클레오디드가 결합된 긴 사슬의 상보적 결합 알파 나선이라고도 함
다양한 형태의 DNA존재

34
여러가지 형태의 이중나선

35
DNA 분자구조의 특징 Highly regular fashion duplex(helical chain)
A-T & C-G (Chagaffe's law)
")
37
사람의 nucleotide의 수: 약 32억 b.p.
생성가능한 변이의 수:432억 길이: 3x106개의 nucleotide가 1mm 따라서 30억/300백만=1000mm=1m

38
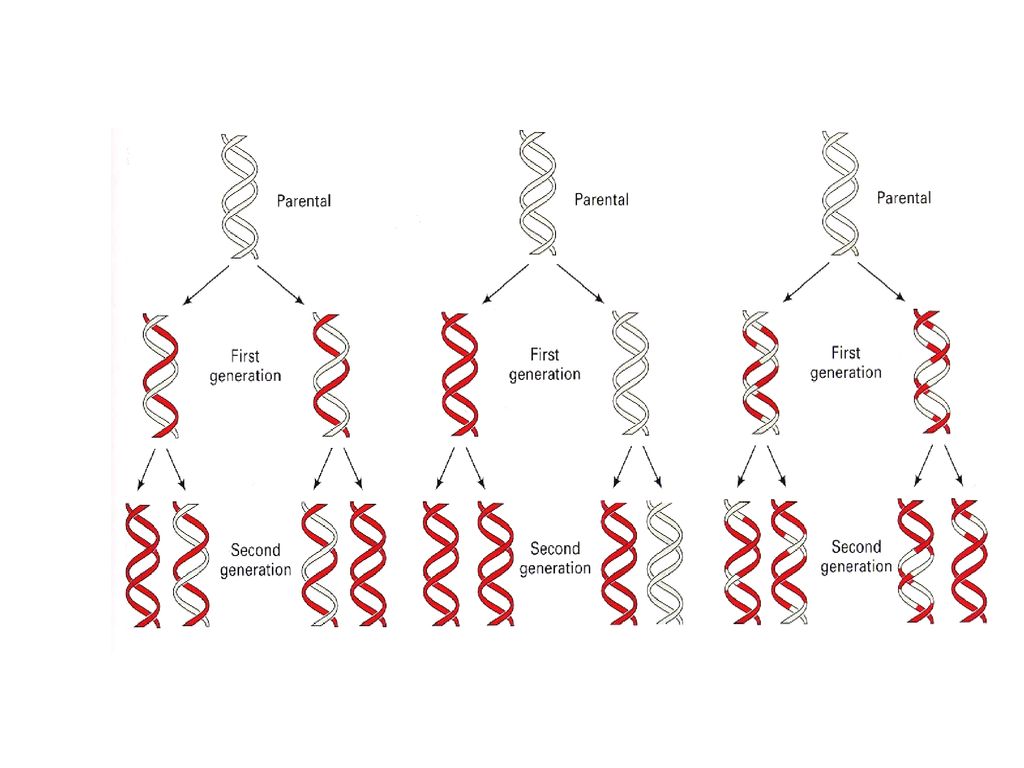
DNA replication

42
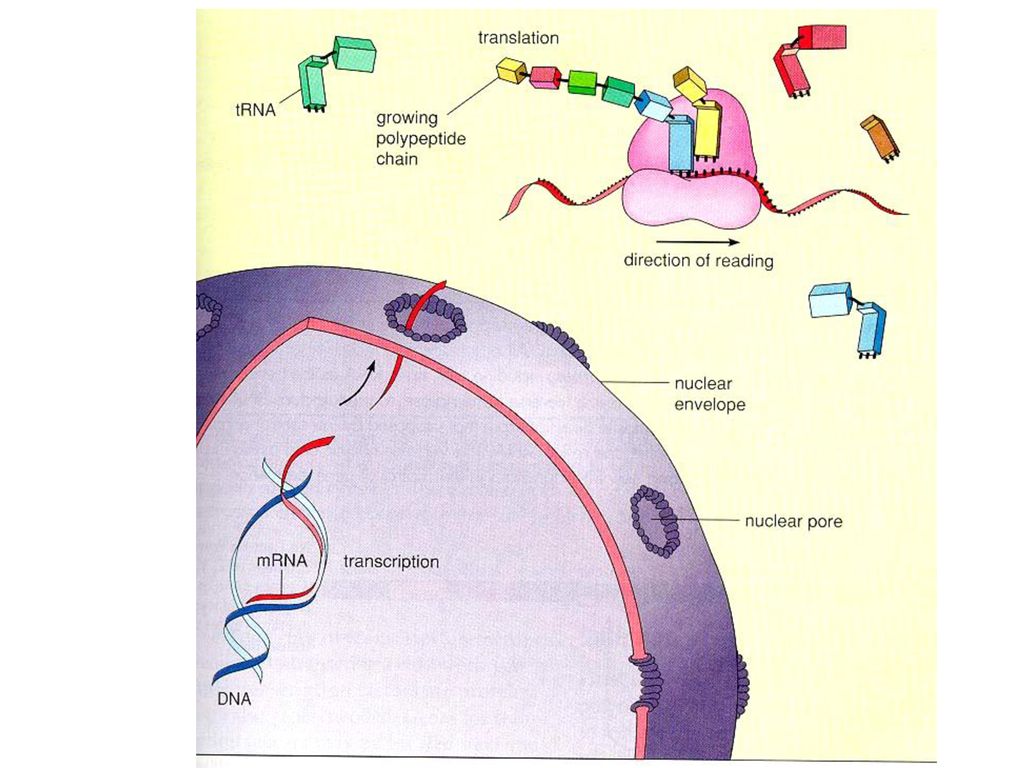
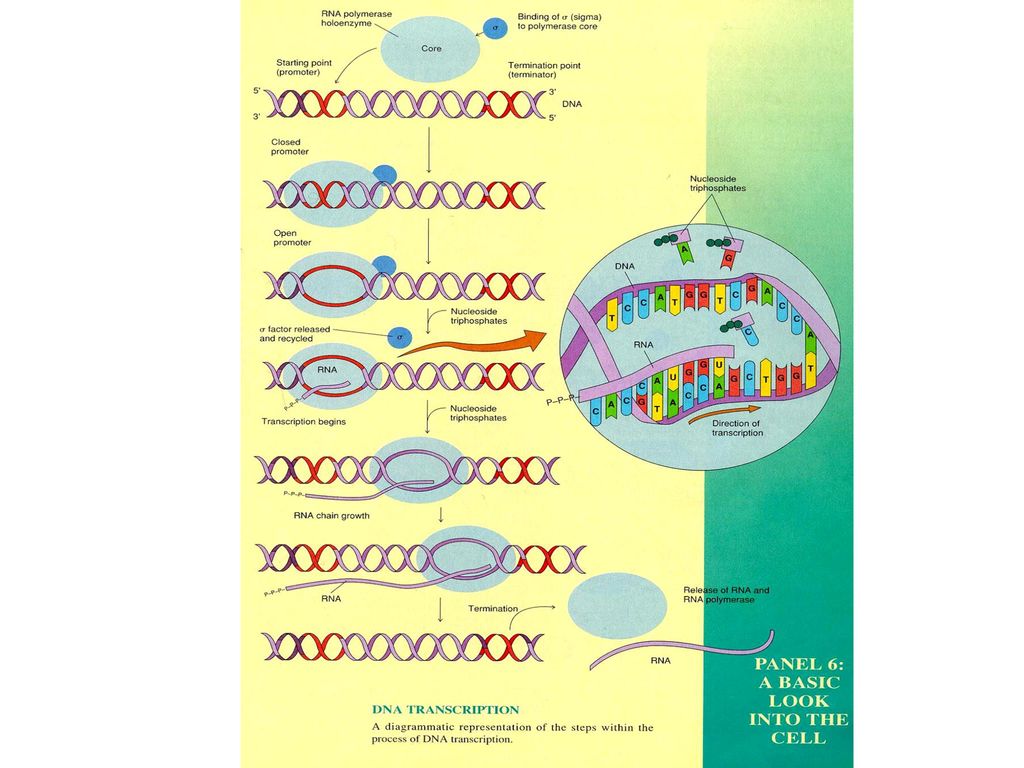
Transcription

43
Central dogma DNA > mRNA > Amino Acid transcription translation the kinds of RNA mRNA tRNA rRNA

46
mRNA synthesis 5'--> 3' RNA sequence는 DNA에 의해 결정
DNA double strand 중 한 strand만 copy (non-sense strand & sense strand) DNA의 Thymine대신 Uracil Processing o binding o initiation: AUG codon o elongation o termination: UGA, UAG or UAA
 DNA의 Thymine대신 Uracil. Processing. o binding. o initiation: AUG codon. o elongation. o termination: UGA, UAG or UAA.")
48
18.단백질 합성

49
목차 1. 유전암호 ① 코돈안티코돈의 상호작용 2. 단백질 합성 ① 원핵생물의 단백질 합성 ②아미노아실- tRNA 합성요소
② 진핵세포의 단백질 합성 ③Folding의 문제점

50
단백질 합성 단백질의 종류에 따라 형질이 달라짐 :체내의 형질은 DNA의 유전정보에 따라 달라짐
형질의 발현과정= 단백질 합성과정 1. 코드(DNA) : 유전정보 전사(복사) 2. 코돈(mRNA) : 전사된 유전정보 운반 3. 안티코돈(tRNA) : 번역과 아미노산의 배열 4. 단백질 합성( 특정 형질 발현)
 : 유전정보 전사(복사) 2. 코돈(mRNA) : 전사된 유전정보 운반. 3. 안티코돈(tRNA) : 번역과 아미노산의 배열. 4. 단백질 합성( 특정 형질 발현)")
51
단백질 합성 리보솜

52
리보솜

53
단백질 합성 ◎리보솜,mRNA, tRNA, 그리고 많은 단백질 인자를 필요로 하는 복잡한 과정
① 리보솜<ribosome> 단백질이 합성되는 장소 ② mRNA DNA의 coding 결과로부터 RNA 중합효소에 의해 만들어지고,messenger역할 ③ tRNA 아미노산을 리보솜으로 운반

54
유전암호(genetic code) ◎아데닌<adenine> C5H5N5 아미노퓨린구조를 가지는 염기성물질
◎ 시토신<cytosine> 옥시-6-아미노피리미딘 ◎ 구아닌<guanine> C5H5N5O 2-아미노-6-옥시퓨린에 해당하는 핵산 구성성분인 퓨린 염기의 일종. ◎ 우라실<uracil> C4H4N2O2 생물계에 널리 존재하는 피리미딘 염기의 유도체. RNA 속에 함유되어 있다.

55
코돈(codon) ◎ DNA를 전사하는 mRNA의 3염기 조합 즉, mRNA의 유전암호의 단위
①개시코돈(initiation codon) 염기배열이 유전정보로서 올바르게 번역되려면, 어떤 특정한 위치에서 번역이 개시될 필요가 있는데, 이때 개시점이 되는 코돈 ② 종결코돈(termination codon) 번역을 미리 종결시켜 버려 암호화 부위의 정상적인 말단과 매우 근접해 있지 않는 한 아무런 기능을 못하는 폴리펩티드를 만드는코돈
 염기배열이 유전정보로서 올바르게 번역되려면, 어떤 특정한 위치에서 번역이 개시될 필요가 있는데, 이때 개시점이 되는 코돈. ② 종결코돈(termination codon) 번역을 미리 종결시켜 버려 암호화 부위의 정상적인 말단과 매우 근접해 있지 않는 한 아무런 기능을 못하는 폴리펩티드를 만드는코돈.")
56
유전암호의 특성 1.퇴행성 코돈 (Degenerative codon) 아미노산을 암호화하는 코돈이 한 개 이상 있는 것을 말한다. 2.특이성 (Specificity) 각 코돈은 특정한 아미노산을 암호화하는 부호인데 같은 아미노산을 암호화하는 대부분의 코돈은 매우 유사한 염기서열을 갖고 있다. 3.비중첩 암호 (Nonoverlapping codon) mRNA 코딩서열은 종결코돈에 도달 할때까지 한번에 3개의 염기를 연속해서 리보솜이 읽어간다. 4.보편성 (Universality) 대부분의 유전암호는 보편성을 띠고 있다. 해독과정 조사 결과 아미노산을 코딩 하는 부호는 항상 같다.
 각 코돈은 특정한 아미노산을 암호화하는 부호인데 같은 아미노산을 암호화하는 대부분의 코돈은 매우 유사한 염기서열을 갖고 있다. 3.비중첩 암호 (Nonoverlapping codon) mRNA 코딩서열은 종결코돈에 도달 할때까지 한번에 3개의 염기를 연속해서 리보솜이 읽어간다. 4.보편성 (Universality) 대부분의 유전암호는 보편성을 띠고 있다. 해독과정 조사 결과 아미노산을 코딩 하는 부호는 항상 같다.")
57
코돈-안티코돈 상호작용(Codon-anticodon interaction)
◎유전정보를 번역하는 과정에서 mRNA상의 코돈과 짝이 되는 안티코돈이 코돈과 아미노산의 사이를 특이적으로 대응 코돈과 안티코돈의 염기쌍 UGC코돈 GCA안티코돈 짝을 이루어야 cys-teine 아미노산이 폴리펩티드를 만들 수 있다.

58
동요가설(wobble hypothesis)
◎ 세포가 예상된 기대치보다 적은 숫자의 tRNA를 갖고 단백질을 합성할 수 있는가를 설명해주는 학설 (예상치 61개보다 적은 약 50개의 tRNA 를 갖고 있다.) ① 코돈과 안티코돈 상호작용에 있어 첫 두 염기쌍이 단백질 해독 중 필요한 특이성을 제공 ②세 번째 코돈과 안티코돈 염기 사이의 상호작용과 결합은 덜 엄격 즉,비정통적인 염기 쌍이 가끔 나타남.
 ① 코돈과 안티코돈 상호작용에 있어 첫 두 염기쌍이 단백질 해독 중. 필요한 특이성을 제공. ②세 번째 코돈과 안티코돈 염기 사이의 상호작용과 결합은 덜 엄격 즉,비정통적인 염기 쌍이 가끔 나타남.")
59
아미노아실-Trna 합성효소(Aminoacyl-Trna synthetase)
◎활성화반응(Activation) 혼합 무수물 결합을 형성함으로 아미노산을 활성화시키는 반응은 ATP의 가수분해로 일어나고 생성물은 피로인산(pyrophosphate) ◎ tRNA dusruf (linkage) 합성효소의 활성부위에 결합된 특정 tRNA은 에스테르 결합을 통해 아미노아실기에 결합 아미노아실-tRNA의 형성
 혼합 무수물 결합을 형성함으로 아미노산을 활성화시키는 반응은 ATP의 가수분해로 일어나고 생성물은 피로인산(pyrophosphate) ◎ tRNA dusruf (linkage) 합성효소의 활성부위에 결합된 특정 tRNA은 에스테르 결합을 통해 아미노아실기에 결합. 아미노아실-tRNA의 형성.")
60
단백질의 합성 3단계 1) 개시 (initiation) 2) 연장 (elongation) 3) 종결 (termination)
 개시 (initiation) 2) 연장 (elongation) 3) 종결 (termination)")
61
단 백 질 의 합 성

62
1) 개시 리보솜 소단위 와 mRNA결합시 발생 (acyl site)존재. ◎ 해독(translation)의 개시
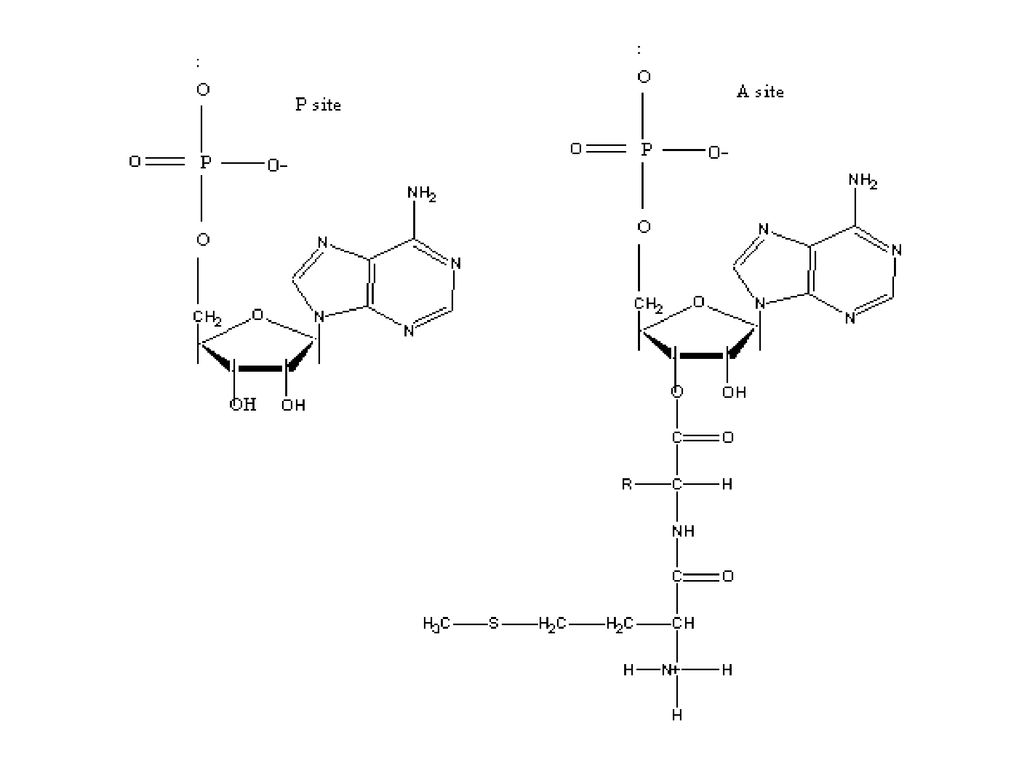
◎ 개시 tRNA의 안티코돈은 개시코돈AUG와 염기쌍을 이룸. ◎ 리보솜: P부위(peptide site)와 A부위 (acyl site)존재. ◎ 리보솜+리보솜+….+리보솜=폴리솜
와 A부위. (acyl site)존재. ◎ 리보솜+리보솜+….+리보솜=폴리솜.")
63
2) 연장 ◎ 폴리펩티드 합성 (촉매: 펩티드 전이효소) ◎ 펩티드 결합의 연장주기는 종결 코돈이A부위에 들어올 때까지 반복.
N-말단으로부터 C-말단으로 진행 ◎ 펩티드 결합의 연장주기는 종결 코돈이A부위에 들어올 때까지 반복.

64
<펩티드 결합의 합성>

66
3) 종결 ◎ 폴리펩타이드: 리보솜으로부터 방출 ◎ 종결코돈과 아미노아실-tRNA의 결합불 가능 해독 종결 단백질 방출인자는 A부위에 결합
 종결 ◎ 폴리펩타이드: 리보솜으로부터 방출 ◎ 종결코돈과 아미노아실-tRNA의 결합불 가능 해독 종결 단백질 방출인자는 A부위에 결합")
67
4)단백질의 합성<리보솜에서>
단백질의 합성<리보솜에서>")
68
< 단백질 합성 과정 요약 > 1. 핵 속에서 DNA 이중 나선이 풀리면서 → mRNA가 전사 2. 전사된 mRNA는 핵공을 통해 세포질로 나와 → 리보솜(30S)과 결합 3. mRNA의 코돈에 대응하는 아미노산을 → tRNA 가 리보솜(50S)로 운반 4. 운반된 아미노산을 펩티드 결합하여 → 폴리펩티드 사슬로 신장 5.단백질로 합성되어 효소나 호르몬, 항체, 헤모글로빈 등으로 형질이 발현됨
과 결합 3. mRNA의 코돈에 대응하는 아미노산을 → tRNA 가 리보솜(50S)로 운반 4. 운반된 아미노산을 펩티드 결합하여 → 폴리펩티드 사슬로 신장 5.단백질로 합성되어 효소나 호르몬, 항체, 헤모글로빈 등으로 형질이 발현됨")
69
< 단백질 합성 과정 요약 >

70
원핵생물의 단백질합성 -단세포 -29∼34억 년 전 발생추정. -생물진화에서 가장 오래된 것으로 추정.
◎ 원핵생물이란? -단세포 -29∼34억 년 전 발생추정. -생물진화에서 가장 오래된 것으로 추정. -분자 상태로 세포질 내에 존재. -미토콘드리아등의 구조체가 없음. -해독은 비교적 빠름.

71
1)개시 2)연장 ①원핵 생물은 세포질에서 RNA전사와 단백질 합성이 같이 일어남. ②샤인-달가노 서열
①아미노아실-tRNA를 A부위에 위치. ②펩티드 결합 형성 ③자리 옮김(translocation)
")
72
<원핵세포의 개시복합체 형성>

73
<대장균의 EF-Tu-EF-ts의 주기>

74
3)종결 ①종결 코돈(UAA,UAG,UGA)이 A부위에 들어갈 때 시작. ②폴리펩타이드 리보솜에서 방출
mRNA와 tRNA로 분리. ③리보솜이 소단위로 분리 될 때 끝남.

75
4)해독 후 변형 ①단백질 가수분해 가공: 단백질 가수분해 작용에 의해 포르밀메티오닌 잔기, 신호펩티드등을 제거하는 절단반응 발생. ②글리코실화반응: 소수의 당단백질이 세포표면에 존재. ③메틸화 반응: 메틸전이효소가 S-아데노실메티오닌을 이용하여 메틸화반응. ④인산화 반응: 단백질 키나아제 이용

76
5)해독 통제 기작 ①단백질 합성 에너지 소모 多 펩티드결합1개,고에너지 인산결합 필요. ②전사와 해독과정 직접 연결
③원핵세포 mRNA의 반감기(1~3분)는 비교적 짧다.
는 비교적 짧다.")
77
<대장구균의 전사와 해독> a)대장균전사와 해독을 찍은 전자현미경 사진
대장균전사와 해독을 찍은 전자현미경 사진")
78
b)해독과정의 폴리리보솜의 모습
해독과정의 폴리리보솜의 모습")
79
<음성 해독 조절>

80
<음성 해독 조절> ①대장균의 PL11오페론은 농도에의해 통제. ②리보솜단백질 L1은 23S rRNA또는 mRNA와 결합 가능. ③과다한 농도의 L1은 PL11 mRNA의 해독 억제.

81
진핵세포의 단백질 합성 ◎ 진핵세포의 단백질 합성 (Eucaryotic protein systhesis) ◎단백질 합성순서
원핵세포보다 더 느리고 복잡하다. ◎단백질 합성순서 개시 연장 종결 해독 후 변형 해독 통제 기작

82
◎ 개시 (initiation) mRNA 2차구조 ① 여러 가공 반응을 통해 2차구조 형성
② 단백질과 결합하여 ribonucleoprotein 복합체 생성 ③해독되기 전 2차 구조 변형, 단백질 제거 2. mRNA 주사(scanning) ① 리보솜 capped 5’-end ② 5’ ’방향으로 이동 결 합
 ① 리보솜 capped 5’-end. ② 5’ 3’방향으로 이동. 결 합.")
83
◎진핵세포의 개시는 작은 40S는리보솜소단위가 eEF2 (GTP 결합단백질), GTP와 개시 metyionyl-tRNAmet (met-tRNAi)으로 구성된 복 합체와 결합할 때 시작한다.
으로 구성된 복. 합체와 결합할 때 시작한다.")
84
◎진핵생물에서 개시 tRNA는 처음에 개시인자와 함께 리보솜의 작은 소 단위체에 적재된다. 많은 tRNA중에 개시tRNA만이
리보솜의 작은 소단위체에 단단하게 결합될 수 있다. ◎ 개시tRNA를 적재한 리보솜의 작은 소단위체는 진핵생물 mRNA에 있는 cap을 부분적으로 인식하여 mRNA 분자의 5' 말단에 결합한다. ◎ 작은 소단위체가 개시코돈인 AUG를 찾기 위해 mRNA를 따라 5'→3'방향 으로 움직인다.

85
소단위체로부터 떨어지고 큰 소단위체 가 결합하여 리보솜을 완성한다. ◎ 두 번째의 tRNA가 리보솜의 A 부위에
◎ AUG코돈을 만나면 개시인자들이 작은 소단위체로부터 떨어지고 큰 소단위체 가 결합하여 리보솜을 완성한다. ◎ 두 번째의 tRNA가 리보솜의 A 부위에 결합하여 첫 번째 아미노산인 메티오닌 과 2번 째 아미노산을 연결시킨다. ◎ 시작tRNA는 메티오닌을 다음 아미노산 과 연결시켜주고 자신은 분리되어 E부위 로 이동한다음 리보솜으로부터 분리되고, 두 번째 tRNA는 메티오닌과 두 번째 아미 노산을 부착한 채로 P부위로 옮겨 간다.

86
◎ 연장 ①A site에 아미노아실 tRNA 결합 ② 펩티드결합 전이 ③ 자리옮김(translocation)
② 펩티드결합 전이 ③ 자리옮김(translocation) 연장과정이 끝나게 되면… 리보솜 A 부위가 비게 되어 새로운 코돈이 들어오게 된다.
 연장과정이 끝나게 되면… 리보솜 A 부위가 비게 되어. 새로운 코돈이 들어오게 된다.")
87
◎ 종결 ①단백질합성의 종결: 종결코돈(stop codon) 이라 불리는 세 개의 코돈(UAA, UAG, UGA)
중 한 개만 있으면 된다. 이들 코돈은 tRNA에 의해 인지되지 않고 아미노산을 지정하지도 않지만, 그 대신 리보솜에게 번역을 멈추도록 알린다. ②mRNA의 종결코돈이 리보솜의 A부위에 들어오면, 방출인자(release factor)라는 단백질이 리보솜의 A부위에 들어와 종결코돈과 결합한다.
라는 단백질이 리보솜의 A부위에 들어와 종결코돈과 결합한다.")
88
③방출인자의 결합은 리보솜에서 펩티딜전달 효소의 활성을 변경하여 아미노산이 아닌 물 분자가 펩티딜 tRNA에 결합하도록 촉매 한다. ④리보솜은 mRNA를 방출하고, 다른 mRNA에서 새로운 단백질합성을 시작할 수 있는 두 개의 소단위체로 분리된다.

89
◎ 해독 후 변형 ①여러 종류의 공유결합에 의한 구조적 변형 ②단백질 합성 중 또는 후에 일어남.
③특정 아미노산의 측쇄(side chain)를 변형시키거나 특정결합 절단 ④해독 후 변형은 단백질의 구조와 기능에 영향을 끼침.
를. 변형시키거나 특정결합 절단. ④해독 후 변형은 단백질의 구조와 기능에. 영향을 끼침.")
90
< 대표적인 해독 후 변형의 예 > 1. 단백질 가수분해에 의한 절단
2. 글리코실화 반응(Glycosylation) 3. 수산화 반응(Hydroxylation) 4. 인산화 반응 5. 친유성 변형 6. 메틸화 반응 7. 이황화 결합 (Disulfide bond) < 프롤린의 수산화반응 >
 3. 수산화 반응(Hydroxylation) 4. 인산화 반응. 5. 친유성 변형. 6. 메틸화 반응. 7. 이황화 결합. (Disulfide bond) < 프롤린의 수산화반응 >")
91
◎ 해독 통제 기작 1. mRNA 운반 ②에너지와 5’-cap과 poly A tail을 갖고 있어야 함. 2. mRNA 안정성
② 3’말단의 가역적인 아데닐 반응, 탈아데닐 반응에 영향 받음. 3. 음성 해독 통제(Negative translation control) ① 5’말단 염기서열에 억제단백질을 결합시킴. 4. 개시인자 인산화(Initiation factor phosphorylation) ① 단백질 합성으로 보통 감소시킴. 5. 해독틀 이동(Translational frameshifting) ① 활성화될 경우 +1또는 -1의 해독틀의 변화 발생
 ① 5’말단 염기서열에 억제단백질을 결합시킴. 4. 개시인자 인산화(Initiation factor phosphorylation) ① 단백질 합성으로 보통 감소시킴. 5. 해독틀 이동(Translational frameshifting) ① 활성화될 경우 +1또는 -1의 해독틀의 변화 발생.")
92
◎ Folding의 문제점 8M 요소와 메르캅탄(mercaptan, RSH)에 의해 변성화된 RNA 가수분
해효소가 메르캅탄을 제거함으로써 원래 상태의 활성화된 상태로 전환됨. (크리스티안 안핀센) 아미노산의 물리적, 화학적 성질과 폴딩과정에 관련된 결합회전, 자유에너지 등에 대한 정보를 알면 단백질의 3차구조를 예측할 수 있음.
 아미노산의 물리적, 화학적 성질과 폴딩과정에 관련된 결합회전, 자유에너지 등에 대한 정보를 알면 단백질의 3차구조를 예측할 수 있음.")
93
◎ 단백질 folding에 중요한 요소 ◎ 단백질 folding과 표적화 ①단백질의 2차구조(α-나선과 β-병풍구조)
②소수성 결합(hydrophobic interaction) ◎ 단백질 folding과 표적화 분자 샤페론(molecular chaperone)이라고 하는 분자들에 의해 조성되는 것이 밝혀짐.
 ◎ 단백질 folding과 표적화. 분자 샤페론(molecular chaperone)이라고 하는. 분자들에 의해 조성되는 것이 밝혀짐.")
94
◎ 샤페론 1. Hsp 70 ①folding 초기에 단백질을 안정시킴. ②단백질의 소수성 부위에 결합하여 응집 방해
폴리펩티드를 최종적으로 folding 시킴.

95
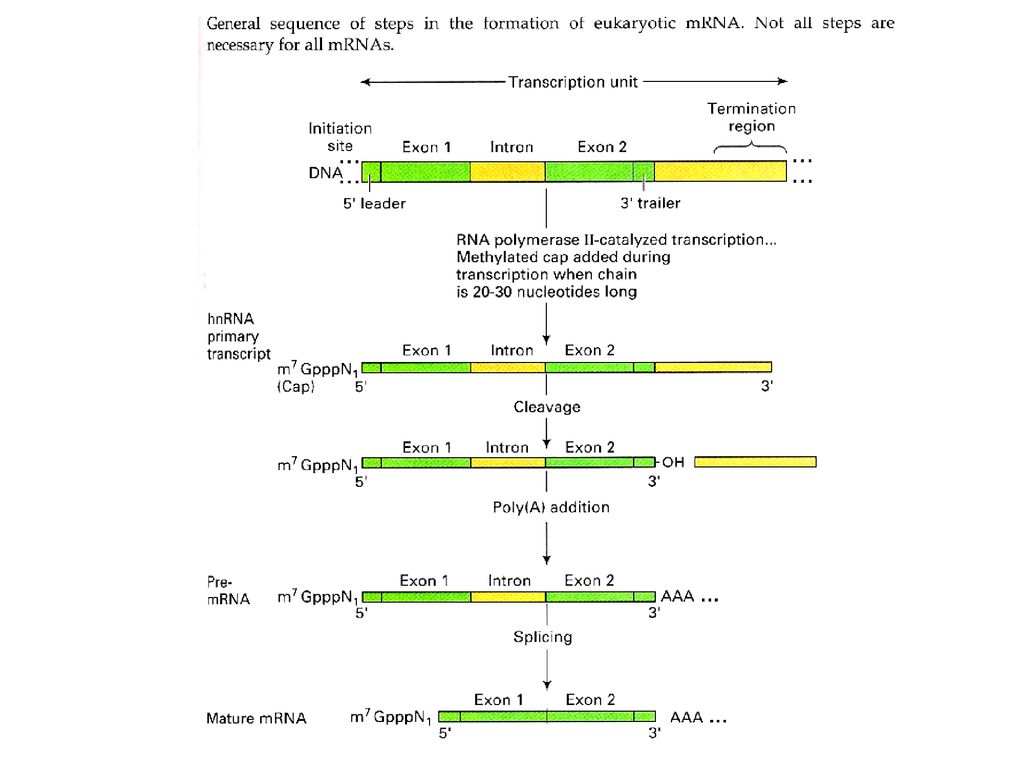
Post-transcription processing
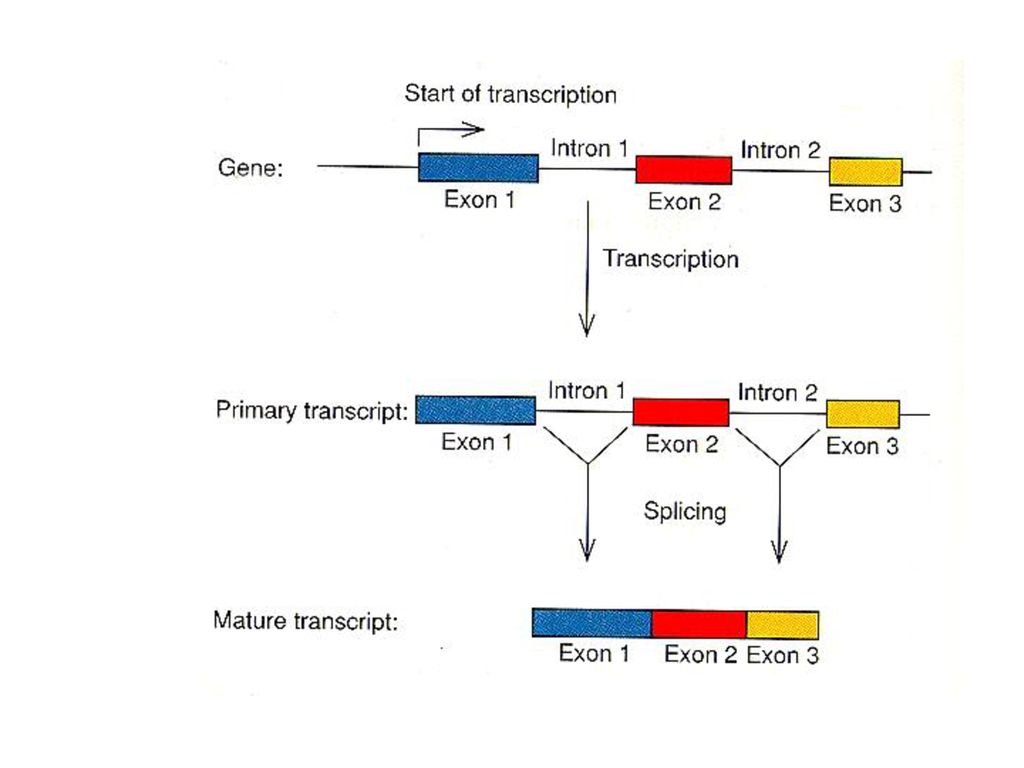
Pre-mRNA Capping 5'-m7G ppp----3'OH - 다른 nucleotide의 부착방지 - capping 부분이 적합한 ribosome 인지 Poly A tail 5'------AAAA--AA3'OH RNA excision & splicing exon: coding seq. intron: non-coding seq.

98
tRNA transcription & processing
anticodon이 mRNA codon과 결합 한 종류의 tRNA가 여러종류의 mRNA와 binding anticodon 5' position에 inosine(I) base를 가지기도 한다.(inosine은 adenine이 변형된 형태) wobble hypothesis 한 amino acid를 결정하는 codon이 몇개의 규칙적인 codon으로 되어있음을 설명하는 것으로 1개의 amino acid에 대하여 첫 2 base sequence는 같으나 3번째 base가 vary하다는 것이다.
 base를 가지기도 한다.(inosine은 adenine이 변형된 형태) wobble hypothesis. 한 amino acid를 결정하는 codon이 몇개의 규칙적인 codon으로 되어있음을 설명하는 것으로 1개의 amino acid에 대하여 첫 2 base sequence는 같으나 3번째 base가 vary하다는 것이다.")
103
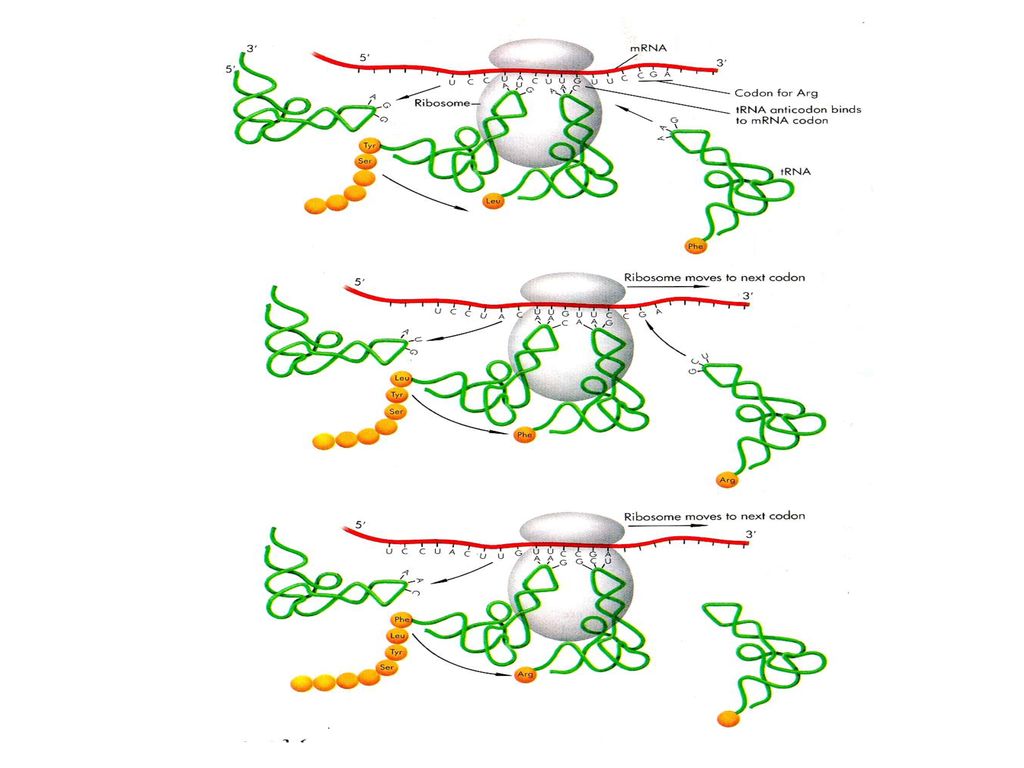
Translation













 전기영동 (Electrophoresis) 전하를 띤 고분자 물질을 전기장을 띤 매질 ( 젤 ) 에서 이동 ∙ 분리 물질의 분리, 순도 ∙ 특성 분석 물질의 이동.>")



: 화학적인 방법으로는 더 이상 간단한 물질로 나눌 수 없는 물질을 말함. 현재 지구상에는 92 가지의 자연 원소가 존재 SPONCH: 황 (sulfur), 인 (phosphorus), 산소.>")










 - 효소의 중요성 - 대사 경로 (catabolism) - 세포구성분의 생합성 (Anabolism)>")

.>")
와 물(H2O)을 탄수화물과 같은 유기화합물로 전환시키고 산소(O2)를 방출하는 복잡한 과정으로서 명반응(明反應)과 암반응(暗反應)으로 구분할 수 있다. 명반응은 엽록체의.>")
이란? 게놈(genome)은 유전자(gene)와 염색체(chromosome)>")