Download presentation
1
아이스 웜(ice worm)인 Hesiocaeca methanicola은 그 어떤 광원이나 광합성생물로부터 멀리 떨어져 있는 멕시코 만 바닥의 메탄 아이스(methane ice)에서 자라고 있는 세균을 먹고 산다.
인 Hesiocaeca methanicola은 그 어떤 광원이나 광합성생물로부터 멀리 떨어져 있는 멕시코 만 바닥의 메탄 아이스(methane ice)에서 자라고 있는 세균을 먹고 산다.")
2
생물은 광합성에 의존한다. 광합성 (photosynthesis): 태양에너지가 특정 분자들의 결합에 의해 포도당의 생성에 이용되는 과정. 반응물은 CO2와 H2O이며, 생성물은 포도당 (C6H12O6), H2O, O2이다. 6CO2+12H2O → C6H12O6 + 6O2 + 6H2O (△G + 686kcal/mole ; 흡열반응) 이 때 발생하는 산소 (O2) 는 CO2가 아니라 H2O에서 나온 것이다. 일부 미생물의 경우 H2O 대신에 H2S 를 사용하여 광합성을 한다. 이 경우 산소 (O2) 대신에 황 (S2) 를 배출한다.
 이 때 발생하는 산소 (O2) 는 CO2가 아니라 H2O에서 나온 것이다. 일부 미생물의 경우 H2O 대신에 H2S 를 사용하여 광합성을 한다. 이 경우 산소 (O2) 대신에 황 (S2) 를 배출한다.")
4
광합성은 어떻게 진화되었나? 그림 4.2 산소는 생명 세계를 변화시켰다.
대기의 산소가 항상 풍족했던 것은 아니다. 광합성 진화는 산소를 대기로 펌프질하여, 생명체의 다양성을 근본적으로 변화시켰다. 그림 4.2 산소는 생명 세계를 변화시켰다. 대기의 산소가 항상 풍족했던 것은 아니다. 광합성 진화는 산소를 대기로 펌프질하여, 생명체의 다양성을 근본적으로 변화시켰다.

6
Isaac Newton: 백색광은 보라색부터 붉은색 단색광의 혼합물이다. James Maxwell (1860):
가시광선은 전자기파 (Electromagnetic waves) 의 일부분이다. Albert Einstein (1905): 빛은 광자 (photon) 의 단위로 되어 있다. 짧은 파장이 고에너지. 태양빛: 4% 자외선, 52% 적외선, 44% 가시광선 대기의 산소가 항상 풍족했던 것은 아니다. 광합성 진화는 산소를 대기로 펌프 질하여, 생명체의 다양성을 근본적으로 변화시켰다.
 의 일부분이다. Albert Einstein (1905): 빛은 광자 (photon) 의 단위로 되어 있다. 짧은 파장이 고에너지. 태양빛: 4% 자외선, 52% 적외선, 44% 가시광선. 대기의 산소가 항상 풍족했던 것은 아니다. 광합성 진화는 산소를 대기로 펌프. 질하여, 생명체의 다양성을 근본적으로 변화시켰다.")
7
광합성 세균을 제외한 대부분의 식물, 조류의 주된 광합성 색소
엽록소 a (chlorophyll a): 광합성 세균을 제외한 대부분의 식물, 조류의 주된 광합성 색소 붉은색과 주황색 ( nm), 파란색과 보라색 ( nm) 흡수 녹색 파장 반사 그림 4.4 잎의 색. 잎은 잎세포의 엽록소분자가 녹색과 노란색 파장은 반사하고 다른 파장은 흡수하기 때문에 녹색으로 나타난다.
: 광합성 세균을 제외한 대부분의 식물, 조류의 주된 광합성 색소. 붉은색과 주황색 ( nm), 파란색과 보라색 ( nm) 흡수. 녹색 파장 반사. 그림 4.4 잎의 색. 잎은 잎세포의 엽록소분자가 녹색과 노란색 파장은 반사하고 다른 파장은 흡수하기 때문에 녹색으로 나타난다.")
8
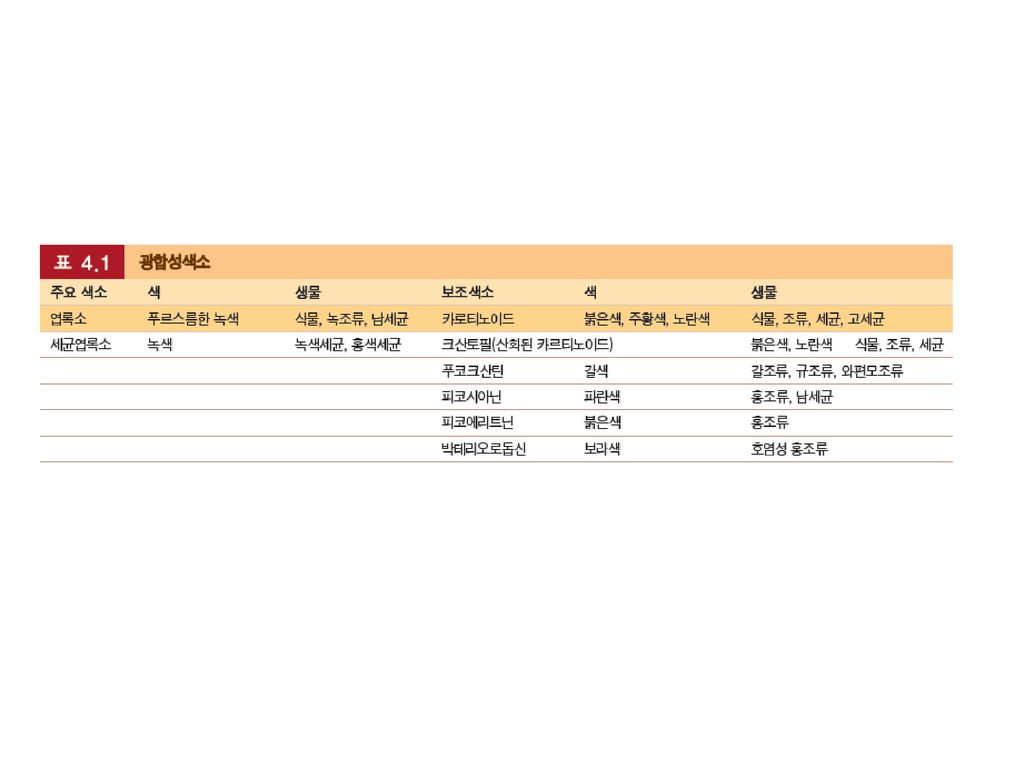
엽록소만큼 많지는 않지만, 보조색소는 엽록소가 흡수하지 못하는 빛의 파장을 흡수한
다. 이러한 색소에는 카로티노이드, 피코에리트린과 피코시아닌이 포함된다. 그림 4.5 다른 광합성색소들. 엽록소만큼 많지는 않지만, 보조색소는 엽록소가 흡수하지 못하는 빛의 파장을 흡수한다. 이러한 색소에는 카로티노이드, 피코에리트린과 피코시아닌이 포함된다.

9
그림 4.6 엽록소. 녹색 색소인 엽록소 a는 식물의 광합성세포의 주요 색소이다. 긴 소수성 꼬리는 분자를 엽록체의 틸라코이드막에 고정시킨다.

11
색소체 (Plastid): 엽록체, 녹말채, 잡색체 세포당 40-200개 존재, 500,000/1mm2
엽록체는 광합성이 일어나는 곳이다. 색소체 (Plastid): 엽록체, 녹말채, 잡색체 세포당 개 존재, 500,000/1mm2 잎의 엽육조직은 많은 엽록체를 갖고 있는 세포로 구 성되어 있다. 막의 이중층은 그라나라 불리는 더미로 조직화되는, 틸라코이드라 불리는 납작하고 서로 연결 되어 있는 주머니를 형성한다. 빛은 흡수된 후 그라나 에서 화학에너지로 전환된다. 화학에너지는 스트로마 에서 탄수화물을 만드는 데 이용된다. 그림 4.7 엽록체: 광합성 장소. 잎의 엽육조직은 많은 엽록체를 갖고 있는 세포로 구성되어 있다. 막의 이중층은 그라나라 불리는 더미로 조직화되는, 틸라코이드라 불리는 납작하고 서로 연결되어 있는 주머니를 형성한다. 빛은 흡수된 후 그라나에서 화학에너지로 전환된다. 화학에너지는 스트로마에서 탄수화물을 만드는 데 이용된다.
: 엽록체, 녹말채, 잡색체. 세포당 개 존재, 500,000/1mm2. 잎의 엽육조직은 많은 엽록체를 갖고 있는 세포로 구. 성되어 있다. 막의 이중층은 그라나라 불리는 더미로. 조직화되는, 틸라코이드라 불리는 납작하고 서로 연결. 되어 있는 주머니를 형성한다. 빛은 흡수된 후 그라나. 에서 화학에너지로 전환된다. 화학에너지는 스트로마. 에서 탄수화물을 만드는 데 이용된다. 그림 4.7 엽록체: 광합성 장소. 잎의 엽육조직은 많은 엽록체를 갖고 있는 세포로 구성되어 있다. 막의 이중층은 그라나라 불리는 더미로 조직화되는, 틸라코이드라 불리는 납작하고 서로 연결되어 있는 주머니를 형성한다. 빛은 흡수된 후 그라나에서 화학에너지로 전환된다. 화학에너지는 스트로마에서 탄수화물을 만드는 데 이용된다.")
12
광계 (photosystem): 광합성을 하는 생명체가 빛에너지의 사용을 가능케 하는 색소분자 덩어리
작용중심 (reaction center): 광합성에서 광에너지를 받아들이는 엽록소 a 와 단백질의 집단 ① 두 광계(two phtosystem) 틸라코이드막에 있는 빛을 포획하고 이용하는 특수구조로 광계I (photosystem I ; PS I)과 광계II (photosystem;PS II) 복합체가 있다. 광수확안테나(light-harvesting antenna)와 반응중심(reaction center)을 가지며, 전자전달계 와 밀접하게 연결 ② 엽록소와 광수확 안테나 광 수확 안테나 : 빛을 포획하여 에너지를 축적하는 일을 하며 엽록소a, 엽록소b (다른 영역 의 파장 흡수), 카로티노이드(carotenoid)로 구성 반응 중심(reaction center) : 에너지가 축적되는 지점으로 빛 에너지가 화학에너지로 전환되는 부위. 한 분자의 엽록소a와 단백질로 구성. 엽록소a를 한 분자씩 갖고 있는 PS I과 PS II 의 반응중심은 각각 700nm 와 680nm 파장 흡수. 엽록소a가 안테나로부터 충분한 에너지를 흡수하면 엽록소의 전자가 들뜨게 되어 궤도에서 이탈 → 엽록소a에서 궤도를 이탈한 전자 는 즉시 에너지를 방출하지 않고 전자 전달계에 포획되어서 화학적인 일을 하는데 사용 (광합성의 특징) 그림 4.8 빛에너지의 수집. 엽록체의 광수확계는 광자를 수집하고 그 에너지를 전자를 통해 작용중심 엽록소로 전달한다.
: 광합성에서 광에너지를 받아들이는 엽록소 a 와 단백질의 집단. ① 두 광계(two phtosystem) 틸라코이드막에 있는 빛을 포획하고 이용하는 특수구조로 광계I (photosystem I ; PS I)과 광계II (photosystem;PS II) 복합체가 있다. 광수확안테나(light-harvesting antenna)와 반응중심(reaction center)을 가지며, 전자전달계 와 밀접하게 연결. ② 엽록소와 광수확 안테나. 광 수확 안테나 : 빛을 포획하여 에너지를 축적하는 일을 하며 엽록소a, 엽록소b (다른 영역 의 파장 흡수), 카로티노이드(carotenoid)로 구성. 반응 중심(reaction center) : 에너지가 축적되는 지점으로 빛 에너지가 화학에너지로 전환되는 부위. 한 분자의 엽록소a와 단백질로 구성. 엽록소a를 한 분자씩 갖고 있는 PS I과 PS II 의 반응중심은 각각 700nm 와 680nm 파장 흡수. 엽록소a가 안테나로부터 충분한 에너지를 흡수하면 엽록소의 전자가 들뜨게 되어 궤도에서 이탈 → 엽록소a에서 궤도를 이탈한 전자 는 즉시 에너지를 방출하지 않고 전자 전달계에 포획되어서 화학적인 일을 하는데 사용 (광합성의 특징) 그림 4.8 빛에너지의 수집. 엽록체의 광수확계는 광자를 수집하고 그 에너지를 전자를 통해 작용중심 엽록소로 전달한다.")
13
광합성의 두 부분 ① 명반응(light reaction) : 빛을 요구하는 반응이며, 계에 에너지를 충전시킴 태양에너지는 ADP와 NADP+를 각각 ATP와 NADPH로 전환시키는 데에 사용 ADP + Pi + NADP+ > ATP +NADPH +H+ ※ 명반응의 생성물은 암반응의 반응물로 사용된다. ② 암반응(dark reaction), 탄소 반응 (carbon reaction) : 빛을 요구하지 않는 반응이며, 계를 원상태로 되돌림 ATP와 NADPH는 포도당 생성에 사용된 후 원래 상태로 되돌아감 CO2 + H2O + ATP + NADPH + H+ -> C6H12O6 + ADP + Pi + NADP+ 틸라코이드막 분자들은 빛에너지를 획득한 후에 에너지를 ATP와 NADPH 분 자로 전달한다. 탄소반응의 효소들은 이 에너지를 이산화탄소를 포획하고 포도 당을 만드는 데 사용한다. 그림 4.9 광합성 개요. 틸라코이드막 분자들은 빛에너지를 획득한 후에 에너지를 ATP와 NADPH 분자로 전달한다. 탄소반응의 효소들은 이 에너지를 이산화탄소를 포획하고 포도당을 만드는 데 사용한다.
, 탄소 반응 (carbon reaction) : 빛을 요구하지 않는 반응이며, 계를 원상태로 되돌림. ATP와 NADPH는 포도당 생성에 사용된 후 원래 상태로 되돌아감. CO2 + H2O + ATP + NADPH + H+ -> C6H12O6 + ADP + Pi + NADP+ 틸라코이드막 분자들은 빛에너지를 획득한 후에 에너지를 ATP와 NADPH 분. 자로 전달한다. 탄소반응의 효소들은 이 에너지를 이산화탄소를 포획하고 포도. 당을 만드는 데 사용한다. 그림 4.9 광합성 개요. 틸라코이드막 분자들은 빛에너지를 획득한 후에 에너지를 ATP와 NADPH 분자로 전달한다. 탄소반응의 효소들은 이 에너지를 이산화탄소를 포획하고 포도당을 만드는 데 사용한다.")
14
그림 4.10 광합성 명반응. 태양에너지는 광계 Ⅱ의 엽록소분자에 의해 포획되고 물 분자로부터 떼어낸 전자에 전달된다. 산소는 부산물로서 방출된다. 에너지가 풍부한 전자는 전자전달계를 구성하는 일련의 운반체를 통해 광계 Ⅰ로 전달된다. 전자가 매번 전달될 때마다 적은 양의 에너지가 제거되어 양성자(물로부터 획득된 수소이온)를틸라코이드 안으로 펌프질하는 데 사용된다. 틸라코이드에 의해 수소이온 농도 기울기가 사라지면 ADP에 인산이 첨가되어 ATP가 형성된다. 광계 Ⅰ에서는 보다 많은 태양에너지가 전자에 더해지고, 전자는 NADP+로 전달되어 에너지가 풍부한 NADPH를 만들어낸다. 에너지 운반체인 ATP와 NADPH는 캘빈회로의탄소반응의 연료로 사용된다. 태양에너지는 광계 Ⅱ의 엽록소분자에 의해 포획되고 물 분자로부터 떼어낸 전 자에 전달된다. 산소는 부산물로서 방출된다. 에너지가 풍부한 전자는 전자전 달계를 구성하는 일련의 운반체를 통해 광계 Ⅰ로 전달된다. 전자가 매번 전달 될 때마다 적은 양의 에너지가 제거되어 양성자(물로부터 획득된 수소이온)를 틸라코이드 안으로 펌프질하는 데 사용된다. 틸라코이드에 의해 수소이온 농도 기울기가 사라지면 ADP에 인산이 첨가되어 ATP가 형성된다. 광계 Ⅰ에서는 보 다 많은 태양에너지가 전자에 더해지고, 전자는 NADP+로 전달되어 에너지가 풍부한 NADPH를 만들어낸다. 에너지 운반체인 ATP와 NADPH는 캘빈회로의 탄소반응의 연료로 사용된다.
를틸라코이드 안으로 펌프질하는 데 사용된다. 틸라코이드에 의해 수소이온 농도 기울기가 사라지면 ADP에 인산이 첨가되어 ATP가 형성된다. 광계 Ⅰ에서는 보다 많은 태양에너지가 전자에 더해지고, 전자는 NADP+로 전달되어 에너지가 풍부한 NADPH를 만들어낸다. 에너지 운반체인 ATP와 NADPH는 캘빈회로의탄소반응의 연료로 사용된다. 태양에너지는 광계 Ⅱ의 엽록소분자에 의해 포획되고 물 분자로부터 떼어낸 전. 자에 전달된다. 산소는 부산물로서 방출된다. 에너지가 풍부한 전자는 전자전. 달계를 구성하는 일련의 운반체를 통해 광계 Ⅰ로 전달된다. 전자가 매번 전달. 될 때마다 적은 양의 에너지가 제거되어 양성자(물로부터 획득된 수소이온)를. 틸라코이드 안으로 펌프질하는 데 사용된다. 틸라코이드에 의해 수소이온 농도. 기울기가 사라지면 ADP에 인산이 첨가되어 ATP가 형성된다. 광계 Ⅰ에서는 보. 다 많은 태양에너지가 전자에 더해지고, 전자는 NADP+로 전달되어 에너지가. 풍부한 NADPH를 만들어낸다. 에너지 운반체인 ATP와 NADPH는 캘빈회로의. 탄소반응의 연료로 사용된다.")
15
그림 4.10 광합성 명반응. 태양에너지는 광계 Ⅱ의 엽록소분자에 의해 포획되고 물 분자로부터 떼어낸 전자에 전달된다. 산소는 부산물로서 방출된다. 에너지가 풍부한 전자는 전자전달계를 구성하는 일련의 운반체를 통해 광계 Ⅰ로 전달된다. 전자가 매번 전달될 때마다 적은 양의 에너지가 제거되어 양성자(물로부터 획득된 수소이온)를틸라코이드 안으로 펌프질하는 데 사용된다. 틸라코이드에 의해 수소이온 농도 기울기가 사라지면 ADP에 인산이 첨가되어 ATP가 형성된다. 광계 Ⅰ에서는 보다 많은 태양에너지가 전자에 더해지고, 전자는 NADP+로 전달되어 에너지가 풍부한 NADPH를 만들어낸다. 에너지 운반체인 ATP와 NADPH는 캘빈회로의탄소반응의 연료로 사용된다. 태양에너지는 광계 Ⅱ의 엽록소분자에 의해 포획되고 물 분자로부터 떼어낸 전 자에 전달된다. 산소는 부산물로서 방출된다. 에너지가 풍부한 전자는 전자전 달계를 구성하는 일련의 운반체를 통해 광계 Ⅰ로 전달된다. 전자가 매번 전달 될 때마다 적은 양의 에너지가 제거되어 양성자(물로부터 획득된 수소이온)를 틸라코이드 안으로 펌프질하는 데 사용된다. 틸라코이드에 의해 수소이온 농도 기울기가 사라지면 ADP에 인산이 첨가되어 ATP가 형성된다. 광계 Ⅰ에서는 보 다 많은 태양에너지가 전자에 더해지고, 전자는 NADP+로 전달되어 에너지가 풍부한 NADPH를 만들어낸다. 에너지 운반체인 ATP와 NADPH는 캘빈회로의 탄소반응의 연료로 사용된다.
를틸라코이드 안으로 펌프질하는 데 사용된다. 틸라코이드에 의해 수소이온 농도 기울기가 사라지면 ADP에 인산이 첨가되어 ATP가 형성된다. 광계 Ⅰ에서는 보다 많은 태양에너지가 전자에 더해지고, 전자는 NADP+로 전달되어 에너지가 풍부한 NADPH를 만들어낸다. 에너지 운반체인 ATP와 NADPH는 캘빈회로의탄소반응의 연료로 사용된다. 태양에너지는 광계 Ⅱ의 엽록소분자에 의해 포획되고 물 분자로부터 떼어낸 전. 자에 전달된다. 산소는 부산물로서 방출된다. 에너지가 풍부한 전자는 전자전. 달계를 구성하는 일련의 운반체를 통해 광계 Ⅰ로 전달된다. 전자가 매번 전달. 될 때마다 적은 양의 에너지가 제거되어 양성자(물로부터 획득된 수소이온)를. 틸라코이드 안으로 펌프질하는 데 사용된다. 틸라코이드에 의해 수소이온 농도. 기울기가 사라지면 ADP에 인산이 첨가되어 ATP가 형성된다. 광계 Ⅰ에서는 보. 다 많은 태양에너지가 전자에 더해지고, 전자는 NADP+로 전달되어 에너지가. 풍부한 NADPH를 만들어낸다. 에너지 운반체인 ATP와 NADPH는 캘빈회로의. 탄소반응의 연료로 사용된다.")
16
그림 4.11 화학삼투적 인산화. 광합성 명반응은 빛에너지를 에너지 운반분자로 전달해준다. 광계 II의 전자는 광자에너지를 흡수하고, 전자전달계를 통해 전달한다. 전자로부터 흡수된 일부 에너지는 수소이온을 틸라코이드 내강으로 펌프질하는 데 사용된다. 전자는 산소를 방출하면서 물로부터 생성된 전자로 대체된다. 광계 II에서 나온 전자는 광계 I로 전달되는데, 전자는 그곳에서 더 많은 에너지를 얻고, 최종적으로 물에서 기인된 수소는 NADP+로 운반되어 NADPH가 만들어진다. ATP 합성효소를 통해 양성자 농도 기울기를 벗어나면서, ADP는ATP로 인산화된다. ATP와 NADPH 모두 스트로마에서 탄소고정을 위한 에너지를 공급해준다. 광합성 명반응은 빛에너지를 에너지 운반분자로 전달해준다. 광계 II의 전자는 광자에너지를 흡수하고, 전자전달계를 통해 전달한다. 전자로부터 흡수된 일부 에너지는 수소이 온을 틸라코이드 내강으로 펌프질하는 데 사용된다. 전자는 산소를 방출하면서 물로부터 생성된 전자로 대체된다. 광계 II에서 나온 전자는 광계 I로 전달되는데, 전자는 그곳 에서 더 많은 에너지를 얻고, 최종적으로 물에서 기인된 수소는 NADP+로 운반되어 NADPH가 만들어진다. ATP 합성효소를 통해 양성자 농도 기울기를 벗어나면서, ADP는 ATP로 인산화된다. ATP와 NADPH 모두 스트로마에서 탄소고정을 위한 에너지를 공급해준다.

17
탄소반응은 탄소를 고정한다 (탄소반응, 암반응) 캘빈회로는 어떻게 탄소를 고정하는가?
6CO2+12H2O → C6H12O6 + 6O2 + 6H2O (△G + 686kcal/mole) 명반응 산물인, ATP와 NADPH는 단순화된 회로로 나타낸 탄소 반응(캘빈회로)을 작동시킨다. 회로는 CO2의 탄소를 유기분자 에 고정시킨다. 루비스코가 첫 단계인 RuBP와 CO2 사이의 반응 을 촉매한다. 탄소반응의 순환은 포도당과 다른 유기 분자를 형 성하고 RuBP를 재생시키는 3탄당 분자인, PGAL를 만든다. 이 경로는 대부분의 광합성 식물에서 작동하지만, 어떤 종은 현저하 게 변형된 회로를 갖고 있다. 그림 4.12 탄소고정. 명반응 산물인, ATP와 NADPH는 단순화된 회로로 나타낸 탄소반응(캘빈회로)을 작동시킨다. 회로는 CO2의 탄소를 유기분자에 고정시킨다. 루비스코가 첫 단계인 RuBP와 CO2 사이의 반응을 촉매한다. 탄소반응의 순환은 포도당과 다른 유기 분자를 형성하고 RuBP를 재생시키는 3탄당 분자인, PGAL를 만든다. 이 경로는 대부분의 광합성 식물에서 작동하지만, 어떤 종은 현저하게 변형된 회로를 갖고 있다.
 명반응 산물인, ATP와 NADPH는 단순화된 회로로 나타낸 탄소. 반응(캘빈회로)을 작동시킨다. 회로는 CO2의 탄소를 유기분자. 에 고정시킨다. 루비스코가 첫 단계인 RuBP와 CO2 사이의 반응. 을 촉매한다. 탄소반응의 순환은 포도당과 다른 유기 분자를 형. 성하고 RuBP를 재생시키는 3탄당 분자인, PGAL를 만든다. 이. 경로는 대부분의 광합성 식물에서 작동하지만, 어떤 종은 현저하. 게 변형된 회로를 갖고 있다. 그림 4.12 탄소고정. 명반응 산물인, ATP와 NADPH는 단순화된 회로로 나타낸 탄소반응(캘빈회로)을 작동시킨다. 회로는 CO2의 탄소를 유기분자에 고정시킨다. 루비스코가 첫 단계인 RuBP와 CO2 사이의 반응을 촉매한다. 탄소반응의 순환은 포도당과 다른 유기 분자를 형성하고 RuBP를 재생시키는 3탄당 분자인, PGAL를 만든다. 이 경로는 대부분의 광합성 식물에서 작동하지만, 어떤 종은 현저하게 변형된 회로를 갖고 있다.")
18
반응1. 카르복시화 반응: CO2가 첨가되는 반응 리블로스이인산 카르복시화 효소(ribulose-1,5-bisphosphate carboxylase ; Rubisco) ⇒ CO2를 리블로스이인산(ribulose 1,5-bisphosphate;RuBp)에 결합시켜 불안정한 6탄당 중간대사물을 형성해 2개의 3탄당으로 분해 PGA (Phospho-Glyceric-Acid) 캘빈회로에서 처음 만들어지는 산물 (C3 plant, C3 pathway) 곡류, 땅콩, 담배, 시금치, 사탕무우, 대두, 잔디.. 반응2. 인산화 : ATP의 고에너지 인산기가 2개의 3탄당 탄수화물로 전이(기질의 자유에너 지 증가) 반응3. 환원 : NADPH가 반응2.의 생성물이 갖고 있는 인산기 중 하나를 수소로 치환 (PGAL) 떨어진 인산은 NADP+및 ADP와 함께 명반응으로 되돌아감
에 결합시켜 불안정한 6탄당. 중간대사물을 형성해 2개의 3탄당으로 분해. PGA (Phospho-Glyceric-Acid) 캘빈회로에서 처음 만들어지는 산물 (C3 plant, C3 pathway) 곡류, 땅콩, 담배, 시금치, 사탕무우, 대두, 잔디.. 반응2. 인산화 : ATP의 고에너지 인산기가 2개의 3탄당 탄수화물로 전이(기질의 자유에너 지 증가) 반응3. 환원 : NADPH가 반응2.의 생성물이 갖고 있는 인산기 중 하나를 수소로 치환 (PGAL) 떨어진 인산은 NADP+및 ADP와 함께 명반응으로 되돌아감.")
19
광합성은 얼마나 효율적일까? 광호흡이 어떻게 캘빈회로에 영향을 주는가? 광호흡 (Photo-respiration)
캘빈회로를 정상적으로 작동시키는 Rubisco의 CO2 포획능력은 CO2가 감소하거나 확산 속도보다 더 빠르게 고정되면 효소의 효율이 현저히 감소 CO2의 농도가 최소 수준 이하로 떨어지면 캘빈회로에서 O2가 CO2를 대체하여 광호흡 (photo-respiration)을 함 산소와 리블로스 이인산이 결합하여 생성된 물질은 여러 단계를 거쳐 글리콜산 (glycolate)이 됨 → 세포질의 미소체(microbody)로 이동해 글리옥실산으로 전환 → 미토콘드리아로 들어가 CO2와 3-PG로 전환 → CO2는 방출되고 3-PG는 연료로 사용
을 함. 산소와 리블로스 이인산이 결합하여 생성된 물질은 여러 단계를 거쳐 글리콜산 (glycolate)이 됨. → 세포질의 미소체(microbody)로 이동해 글리옥실산으로 전환. → 미토콘드리아로 들어가 CO2와 3-PG로 전환. → CO2는 방출되고 3-PG는 연료로 사용.")
20
그림 4.13 C4 광합성. 옥수수 같은 C4 식물의 잎은 명반응과 탄소반응을 물리적으로 분리시키는 독특한 세포구성 (A, B)을 갖고 있다. (C) C4 식물에서, 광합성을 하는 엽육세포는 CO2를 포획하고, 3탄당 화합물인 포스포에놀피루브산(PEP)에 결합시켜 4탄당 화합물인 옥살로아세트산(OAA)을 만든다. OAA는 인접한 다발초세포로 운반되고, 그곳에서 분해되어 CO2와 피루브산을 방출하는 말산과 아스파르트산으로 전환된다. 다발초세포에서의 탄소반응은 방출된 CO2로 포도당을 만들고, 피루브산은 엽육세포로 되돌아간다. 피루브산은 엽육세포에서 PEP로 전환되며 다른 CO2를 취해 C4회로를 다시 시작한다. C4 식물은 다발초세포에서의 CO2 농축으로 광호흡을 감소시킨다. 옥수수 같은 C4 식물의 잎은 명반응과 탄소반응을 물리적으로 분리시키는 독특한 세포구성 (A, B)을 갖고 있다. (C) C4 식물에서, 광합성을 하는 엽육세포는 CO2를 포획하고, 3탄당 화합물인 포스포에놀피루브산(PEP)에 결합시켜 4탄당 화합물인 옥살로아세트산(OAA)을 만든다. OAA 는 인접한 다발초세포로 운반되고, 그곳에서 분해되어 CO2와 피루브산을 방출하는 말산과 아스 파르트산으로 전환된다. 다발초세포에서의 탄소반응은 방출된 CO2로 포도당을 만들고, 피루브 산은 엽육세포로 되돌아간다. 피루브산은 엽육세포에서 PEP로 전환되며 다른 CO2를 취해 C4 회로를 다시 시작한다. C4 식물은 다발초세포에서의 CO2 농축으로 광호흡을 감소시킨다.
을 갖고 있다. (C) C4 식물에서, 광합성을 하는 엽육세포는 CO2를 포획하고, 3탄당 화합물인 포스포에놀피루브산(PEP)에 결합시켜 4탄당 화합물인 옥살로아세트산(OAA)을 만든다. OAA는 인접한 다발초세포로 운반되고, 그곳에서 분해되어 CO2와 피루브산을 방출하는 말산과 아스파르트산으로 전환된다. 다발초세포에서의 탄소반응은 방출된 CO2로 포도당을 만들고, 피루브산은 엽육세포로 되돌아간다. 피루브산은 엽육세포에서 PEP로 전환되며 다른 CO2를 취해 C4회로를 다시 시작한다. C4 식물은 다발초세포에서의 CO2 농축으로 광호흡을 감소시킨다. 옥수수 같은 C4 식물의 잎은 명반응과 탄소반응을 물리적으로 분리시키는 독특한 세포구성 (A, B)을 갖고 있다. (C) C4 식물에서, 광합성을 하는 엽육세포는 CO2를 포획하고, 3탄당 화합물인. 포스포에놀피루브산(PEP)에 결합시켜 4탄당 화합물인 옥살로아세트산(OAA)을 만든다. OAA. 는 인접한 다발초세포로 운반되고, 그곳에서 분해되어 CO2와 피루브산을 방출하는 말산과 아스. 파르트산으로 전환된다. 다발초세포에서의 탄소반응은 방출된 CO2로 포도당을 만들고, 피루브. 산은 엽육세포로 되돌아간다. 피루브산은 엽육세포에서 PEP로 전환되며 다른 CO2를 취해 C4. 회로를 다시 시작한다. C4 식물은 다발초세포에서의 CO2 농축으로 광호흡을 감소시킨다.")
21
C3 식물 : 탄소 고정이 캘빈회로에 의해서만 일어나고 카르복시화에 의한 첫번째 산물이 3탄소 화합물이다.
4탄소 화합물이 관여하는 새로운 CO2 고정 경로 C3 식물 : 탄소 고정이 캘빈회로에 의해서만 일어나고 카르복시화에 의한 첫번째 산물이 3탄소 화합물이다. C4 식물 : CO2는 C3 식물에서와 같이 엽육세포(leaf mesophyll cell)의 엽록체로 확산 PEP카르복시화효소 (phosphoenolpyruvate carboxylase) - CO2 가 처음 만나는 효소로 매우 낮은 CO2 농도에서도 효율적으로 작용 4탄소 대사경로의 최종 산물인 말산(malate)이 엽육세포로부터 인접한 유관속초 세포 (bundle sheath cell ; 엽맥을 에워싸고 있는 세포) 의 엽록체로 이동 CO2 의 농도를 배 높게 유지시킨다. → 4탄소 기질은 3탄소 생성물과 CO2 로 분해 (3탄소 생성물은 엽육세포로 되돌아 가고 CO2 는 Rubisco와 결합하여 갤빈회로로 들어감) C3 식물에 비해 많은 ATP가 요구되나 밝은 조건하에 사는 식물들은 충분한 ATP를 생 성할 수 있다. 옥수수 같은 C4 식물의 잎은 명반응과 탄소반응을 물리적으로 분리시키는 독특한 세포구성 (A, B)을 갖고 있다. (C) C4 식물에서, 광합성을 하는 엽육세포는 CO2를 포획하고, 3탄당 화합물인 포스포에놀피루브산(PEP)에 결합시켜 4탄당 화합물인 옥살로아세트산(OAA)을 만든다. OAA 는 인접한 다발초세포로 운반되고, 그곳에서 분해되어 CO2와 피루브산을 방출하는 말산과 아스 파르트산으로 전환된다. 다발초세포에서의 탄소반응은 방출된 CO2로 포도당을 만들고, 피루브 산은 엽육세포로 되돌아간다. 피루브산은 엽육세포에서 PEP로 전환되며 다른 CO2를 취해 C4 회로를 다시 시작한다. C4 식물은 다발초세포에서의 CO2 농축으로 광호흡을 감소시킨다.
의 엽록체로 확산. PEP카르복시화효소 (phosphoenolpyruvate carboxylase) - CO2 가 처음 만나는 효소로 매우 낮은 CO2 농도에서도 효율적으로 작용. 4탄소 대사경로의 최종 산물인 말산(malate)이 엽육세포로부터 인접한 유관속초 세포. (bundle sheath cell ; 엽맥을 에워싸고 있는 세포) 의 엽록체로 이동 CO2 의 농도를 배 높게 유지시킨다. → 4탄소 기질은 3탄소 생성물과 CO2 로 분해 (3탄소 생성물은 엽육세포로 되돌아 가고. CO2 는 Rubisco와 결합하여 갤빈회로로 들어감) C3 식물에 비해 많은 ATP가 요구되나 밝은 조건하에 사는 식물들은 충분한 ATP를 생 성할 수 있다. 옥수수 같은 C4 식물의 잎은 명반응과 탄소반응을 물리적으로 분리시키는 독특한 세포구성 (A, B)을 갖고 있다. (C) C4 식물에서, 광합성을 하는 엽육세포는 CO2를 포획하고, 3탄당 화합물인. 포스포에놀피루브산(PEP)에 결합시켜 4탄당 화합물인 옥살로아세트산(OAA)을 만든다. OAA. 는 인접한 다발초세포로 운반되고, 그곳에서 분해되어 CO2와 피루브산을 방출하는 말산과 아스. 파르트산으로 전환된다. 다발초세포에서의 탄소반응은 방출된 CO2로 포도당을 만들고, 피루브. 산은 엽육세포로 되돌아간다. 피루브산은 엽육세포에서 PEP로 전환되며 다른 CO2를 취해 C4. 회로를 다시 시작한다. C4 식물은 다발초세포에서의 CO2 농축으로 광호흡을 감소시킨다.")
22
CAM 광합성은 밤에 이산화탄소를 저장한다. 그림 4.14 다른 환경은 광합성에 다른 접근방법을 요구한다.
살고 있는 기후가 워낙 건조하고, 더워서 낮에는 기공을 열 수가 없습니다. 그래서 밤에 기공을 열어서 CO2를 고정 시킨 후에 이것을 저장해 놓았다가 낮에 빛을 받으며 광합성에 이용 합니다. 밤동안에 기공을 열고 PEP carboxylase를 이용해 말산의 형태로 고정시킨 후 이것을 액포에 저장해 놓았다가, 낮에 말산을 피루브산으로 전환시켜서 CO2를 떼어내어 캘빈회로를 진행합니다. CAM 식물은 탄소 고정과 광합성의 진행이 시간적으로 격리되어 있습니다. 그림 4.14 다른 환경은 광합성에 다른 접근방법을 요구한다. 낮은 온도에서, 광호흡은 C3 식물에 부담을 줄 만큼 심각하지 않지만, 높은 온도에서는 광합성을 압도한다. 높은 온도에서, CAM 식물(C)은 광호흡을 감소시키기 위해 밤에 CO2를 저장하지만, C4 식물(B)은 광호흡을 극복하기 위해서 CO2가 높게 농축된 구획을 사용한다. A) 낮은 온도에서, 광호흡은 C3 식물에 부담을 줄 만큼 심각하지 않지 만, 높은 온도에서는 광합성을 압도한다. 높은 온도에서, CAM 식물(C) 은 광호흡을 감소시키기 위해 밤에 CO2를 저장하지만, C4 식물(B)은 광 호흡을 극복하기 위해서 CO2가 높게 농축된 구획을 사용한다.
은 광호흡을 감소시키기 위해 밤에 CO2를 저장하지만, C4 식물(B)은 광호흡을 극복하기 위해서 CO2가 높게 농축된 구획을 사용한다. A) 낮은 온도에서, 광호흡은 C3 식물에 부담을 줄 만큼 심각하지 않지. 만, 높은 온도에서는 광합성을 압도한다. 높은 온도에서, CAM 식물(C) 은 광호흡을 감소시키기 위해 밤에 CO2를 저장하지만, C4 식물(B)은 광. 호흡을 극복하기 위해서 CO2가 높게 농축된 구획을 사용한다.")




 첫번째 행성 지구의 1/3 크기 수성의 1 년 - 88 일 하루 - 59 일 수많은 분화구로 덮여있음 밤낮의 기온차 큼.>")
? 초신성 폭발 원소들은.>")
 와 전자 (-) - 전기적 중성 - 원소의 종류마다 원자핵의 질량과 전자의 개수가 다름.>")
; 태양에너지가 특정 분자들의 결합에 의해 탄수화 물 ( 포도당 ) 의 생성에 이용되는 과정 6CO 2 + 12H 2 OC 6 H 12 O 6 + 6O 2 + 6H 2 O (∆G = +686 kcal/mole)>")







 생태계에서의 에너지 흐름 1. 에너지, 효소와 조절>")
는 변화를 의미. 세포의 모든 화학반응과 물리적인 일에 관련되어 있다.>")






