Download presentation
1
발표 일시: 2013년 11월 4일 월요일 조원: 김연서, 송정현, 강연준, 윤중현, 이정민, 임가은, 이연우, 정다은
생화학 2조 발표 발표 일시: 2013년 11월 4일 월요일 조원: 김연서, 송정현, 강연준, 윤중현, 이정민, 임가은, 이연우, 정다은

2
목 차 Chapter 29 아미노산의 합성 Chapter 30 누클레오티드 대사 Chapter 31 아미노산의 분해와
목 차 Chapter 29 아미노산의 합성 Chapter 30 누클레오티드 대사 Chapter 31 아미노산의 분해와 요소 회로

3
Chapter 29 아미노산의 합성

4
목 차 1. Chapter 29 아미노산의 합성 29.1 질소화효소 복합체는 질소를 고정한다.
목 차 1. Chapter 29 아미노산의 합성 29.1 질소화효소 복합체는 질소를 고정한다. 29.2 아미노산은 주요 대사경로들의 중간물질로부터 만들어진다. 29.3 되먹임 억제(feedback inhibition)는 아미노산 생합성을 조절한다.
는. 아미노산 생합성을 조절한다.")
5
용어 설명 1) 질소(Nitrogen, N) 원소주기율표 상에서 2주기 15족에 속하는 비금속 원소 아미노산의 주요 성분
대기에 풍부 → 지구 대기의 80%를 차지 인체에서의 질소는 단백질, 아미노산, 핵산의 형태로 존재 질소 고정 박테리아로 분류되는 몇몇 박테리아만이 생물권에서 사용 가능한 암모니아의 형태로 질소를 전환시킬 수 있음 ① 글루탐산으로 합성 ② 다른 아미노산 합성에 이용됨

6
N₂+3H₂ 2NH₃ 2) 질소 고정(Nitrogen fixation) 의미 수소 기체와의 혼합을 통해 이루어짐
* 뿌리혹 박테리아 (Rhizobium bacteria) 콩과식물의 뿌리에 침입하여 뿌리혹을 형성 뿌리혹에서 질소가 고정되어 식물과 박테리아에 공급됨 2) 질소 고정(Nitrogen fixation) 의미 질소 고정 미생물(박테리아)들이 생물체가 살 수 있는 조건에서 활성이 없는 질소 기체를 환원하여 두 분자의 암모니아를 생성하는 과정 N₂+3H₂ NH₃ 수소 기체와의 혼합을 통해 이루어짐 철에 의해 촉매 되어 500℃의 온도, 300atm 압력에서 진행됨 (미생물이 극복하기 어려운 환경)
 콩과식물의 뿌리에 침입하여. 뿌리혹을 형성. 뿌리혹에서 질소가 고정되어. 식물과 박테리아에 공급됨. 2) 질소 고정(Nitrogen fixation) 의미. 질소 고정 미생물(박테리아)들이 생물체가. 살 수 있는 조건에서 활성이 없는 질소 기체를. 환원하여 두 분자의 암모니아를 생성하는 과정. N₂+3H₂ 2NH₃. 수소 기체와의 혼합을 통해 이루어짐. 철에 의해 촉매 되어 500℃의 온도, 300atm 압력에서 진행됨. (미생물이 극복하기 어려운 환경)")
7
29.1 질소화효소 복합체는 질소를 고정한다 질소화효소 복합체(Nitrogenase complex)
29.1 질소화효소 복합체는 질소를 고정한다 질소화효소 복합체(Nitrogenase complex) 기본적인 질소 고정 과정을 수행 2개의 단백질로 구성 산소에 민감 → 산소에 의해 비활성화됨 질소 고정은 호기성 생물에게 필수적인 산소에 의해 억제됨 환원효소(Reductase) 높은 환원력을 통해 전자를 제공 질소화효소(Nitrogenase) 환원효소가 제공한 전자를 이용해 질소 기체를 암모니아로 환원시키는 역할을 함
 기본적인 질소 고정 과정을 수행. 2개의 단백질로 구성. 산소에 민감. → 산소에 의해 비활성화됨. 질소 고정은 호기성 생물에게. 필수적인 산소에 의해 억제됨. 환원효소(Reductase) 높은 환원력을 통해 전자를 제공. 질소화효소(Nitrogenase) 환원효소가 제공한 전자를 이용해 질소 기체를. 암모니아로 환원시키는 역할을 함.")
8
1. 질소 기체가 암모니아로 환원되는 과정 N₂+6e-+6H+ 2NH₃ ① 기본적으로 질소 기체가 암모니아로
1. 질소 기체가 암모니아로 환원되는 과정 ① 기본적으로 질소 기체가 암모니아로 환원되는 과정에는 6개의 전자가 관여함 N₂+6e-+6H NH₃

9
N₂+8e-+8H+ 2NH₃+H₂ 8e-=6e-+2e- ② 생물학적 반응 → 항상 질소 기체 1몰당, 최소 1몰의 수소
기체와 2몰의 암모니아가 생성됨 ∴ 추가적으로 2개의 전자 필요 N₂+8e-+8H NH₃+H₂ 8e-=6e-+2e- 추가된 전자 2개

10
N₂+8e-+8H++16ATP+16H₂O 2NH₃+H₂+16ADP+16Pi ③ 전자의 이동과정에서 2분자의 ATP가 가수분해됨
∴ 한 분자의 질소 기체의 환원과정에서 최소한 16분자의 ATP가 가수분해됨 N₂+8e-+8H++16ATP+16H₂O 2NH₃+H₂+16ADP+16Pi

11
질소 고정(Nitrogen fixation)
* ATP 가수분해 전자의 효율적인 이동을 위해 필요한 구조적 변화를 일으킴 반응과정의 에너지 장벽을 낮춰줌 물리적으로 반응과정을 가능하게 함 질소 고정(Nitrogen fixation) 환원된 페레독신에서 오는 전자들 ATP ADP+Pi N₂ * 전자의 흐름 [질소→암모니아] 페레독신 → 환원효소 → 질소화효소 NH₃ 환원효소 (Fe 단백질) 질소화효소 (MoFe 단백질)
 환원된. 페레독신에서 오는 전자들. ATP. ADP+Pi. N₂. * 전자의 흐름. [질소→암모니아] 페레독신 → 환원효소 → 질소화효소. NH₃. 환원효소. (Fe 단백질) 질소화효소. (MoFe 단백질)")
12
2. 질소화효소의 철-몰디브덴 보조인자는 공기 중의 질소와 결합하여 이를 환원 시킨다.
2. 질소화효소의 철-몰디브덴 보조인자는 공기 중의 질소와 결합하여 이를 환원 시킨다. 환원효소와 질소화효소 질소 고정 복합체를 형성 철-황 단백질 전자 전달체 환원효소(Reductase) Iron 단백질 또는 Fe 단백질 동일한 30kd 정도의 단위들이 4Fe-4S 덩어리를 통해 결합한 이량체 구조 적합한 제공자(환원된 페레독신)로부터 전자를 받아 질소화효소로 전자를 전달하는 역할
 Iron 단백질 또는 Fe 단백질. 동일한 30kd 정도의 단위들이 4Fe-4S 덩어리를. 통해 결합한 이량체 구조. 적합한 제공자(환원된 페레독신)로부터 전자를. 받아 질소화효소로 전자를 전달하는 역할.")
13
MoFe 단백질

14
* MoFe 보조인자 * MoFe 단백질 구조 - β subunit(초록색) 2개 → 보조인자란?
2개의 본 ① P 무리 8개 이온 전자 + 7개의 황(6개 시스테인 잔기에 의해 연결) ② MoFe 보조인자 9개의 황 중앙 전자 호모시트르산 (1개의 시스테인, 히스티딘 잔기에 연결) * MoFe 보조인자 → 보조인자란? : 효소에 첨가되는 화학적 인자 - 효소가 활성을 나타내기 위해서 필요한 물질 - 구성성분: 금속이온, 유기물질 질소 고정과정이 일어나는 장소 질소 고정과정: 질소 기체가 암모니아로 전환
 ② MoFe 보조인자. 9개의 황. 중앙 전자. 호모시트르산. (1개의 시스테인, 히스티딘 잔기에 연결) * MoFe 보조인자. → 보조인자란 : 효소에 첨가되는 화학적 인자. - 효소가 활성을 나타내기 위해서 필요한 물질. - 구성성분: 금속이온, 유기물질. 질소 고정과정이 일어나는 장소. 질소 고정과정: 질소 기체가. 암모니아로 전환.")
15
몰리브덴-철-황단백질 Molybdenum-ironsulfur protein 분자상의 N2를 NH3으로 변환하는 질소고정
효소의 2가지 성분 중 한 성분 (다른 성분은 Fe-단백질) 소단위 각2개로 구성하는 사합체(α2β2)를 형성 αβ소단위당 FeMo-보조인자(MoFe2S8)와 P군(2[4Fe-4S])이 1개씩 있음
 소단위 각2개로 구성하는 사합체(α2β2)를 형성. αβ소단위당 FeMo-보조인자(MoFe2S8)와 P군(2[4Fe-4S])이 1개씩 있음.")
16
3. 암모늄 이온은 글루탐산과 글루타민을 통하여 아미노산으로 합성된다.
3. 암모늄 이온은 글루탐산과 글루타민을 통하여 아미노산으로 합성된다. N(질소)+생체분자 NH4+(암모늄 이온)+이용 가능한 아미노산 +: 합성
+생체분자 NH4+(암모늄 이온)+이용 가능한 아미노산 +: 합성.")
17
최소 몇 가지 종에서 NADH, NADPH를 모두 환원제로 사용 가능
* α-케토글루타르산과 글루탐산 암모늄 이온(NH4 +)의 중요한 결합체 α-케토글루타르산: 글루탐산 형성 글루탐산: 글루타민 형성 C O = +NAD(P)H+H + NH4 + + ¯OOC COO¯ α-케토글루타르산 암모늄 이온과 시트르산 회로의 중간 생성물 * 글루탐산 탈수소효소 최소 몇 가지 종에서 NADH, NADPH를 모두 환원제로 사용 가능 +H₃N H 글루탐산 탈수소효소에 의해 반응함으로써 합성됨 C +NAD(P)H+H₂O ¯OOC COO¯ 글루탐산
의 중요한 결합체. α-케토글루타르산: 글루탐산 형성. 글루탐산: 글루타민 형성. C. O. = +NAD(P)H+H + NH4 + + ¯OOC. COO¯ α-케토글루타르산. 암모늄 이온과 시트르산 회로의 중간 생성물. * 글루탐산 탈수소효소. 최소 몇 가지 종에서 NADH, NADPH를 모두 환원제로 사용 가능. +H₃N. H. 글루탐산 탈수소효소에 의해 반응함으로써. 합성됨. C. +NAD(P)H+H₂O. ¯OOC. COO¯ 글루탐산.")
18
- = O O O O O O H +H₃N ¯OOC 글루탐산 ATP H +H₃N 2¯ ADP P ¯OOC 아실-인산 중간체
글루탐산의 곁사슬을 인산화 ¯OOC +H₃N H O = P 2¯ O 아실-인산 중간체 O O

19
= = O O NH₂ O H +H₃N 2¯ P ¯OOC 아실-인산 중간체 NH₃ H +H₃N Pi ¯OOC 글루타민
* 글루타민 질소 제공자 다양한 범위의 중요한 화합물들을 생합성하는 과정에 필요한 질소 원자를 제공 +H₃N H NH₂ 글루타민 ¯OOC = O

20
29.2 아미노산은 주요 대사 경로들의 중간물질로부터 만들어진다
29.2 아미노산은 주요 대사 경로들의 중간물질로부터 만들어진다 아미노산들의 생합성 과정 경로 매우 다양함 공통점 아미노산의 탄소 골격의 공급원이 해당과정, 시트르산 회로, 펜토오스 인산 합성 경로의 중간 생성물들로 한정됨

21
박테리아와 식물에서 아미노산의 생합성 체계 옥살로아세트산 아스팔트산 리신 아스파라진 메티오닌 트레오닌 이소루신 * 표시
파란색 음영: 주요 물질 대사 전구체 노란색: 다른 아미노산을 만들어내는 아미노산 초록색: 필수 아미노산 옥살로아세트산 아스팔트산 리신 아스파라진 메티오닌 트레오닌 이소루신

22
피루브산 리보오스 5-인산 루신 히스티딘 알라닌 발린 포스포에놀피루브산 + 에리트로오스 4-인산 트립토판 페닐알라닌 티로신 티로신

23
α-케토글루타르산 3-포스포글리세르산 글루탐산 세린 글루타민 프롤린 아르기닌 시스테인 글리신

24
1. 인간은 일부 아미노산은 합성할 수 있지만 다른 것들은 반드시 음식물로부터 얻어야 한다
1. 인간은 일부 아미노산은 합성할 수 있지만 다른 것들은 반드시 음식물로부터 얻어야 한다 아미노산의 합성 미생물 : 기본적인 20개의 아미노산을 모두 합성할 수 있음 ex) E. coli 인간 : 11개의 아미노산만을 합성할 수 있음 아미노산: 단백질 분자의 가장 기본적인 조성 물질
 E. coli. 인간. : 11개의 아미노산만을 합성할 수 있음. 아미노산: 단백질 분자의 가장 기본적인. 조성 물질.")
25
필수/비필수 아미노산 합성 경로 단계

26
1) 필수 아미노산 (essential amino acid)
생체 내에서 합성되지 않거나 합성이 어렵기 때문에 음식물로 섭취해야 하는 아미노산 영양분을 통해 반드시 공급되어야 하는 아미노산 종류(성인:8종, 영유아: 9종) 발린(Valine), 루신(Leucine), 아이소루신(Isoleucine), 메티오닌(Methionine), 트레오닌(Threonine), 리신(Lysine), 페닐알라닌(Phenylalanine), 트립토판(Tryptophan) 영유아는 여기에 히스티딘이 더해진다.
 발린(Valine), 루신(Leucine), 아이소루신(Isoleucine), 메티오닌(Methionine), 트레오닌(Threonine), 리신(Lysine), 페닐알라닌(Phenylalanine), 트립토판(Tryptophan) 영유아는 여기에 히스티딘이 더해진다.")
27
필수 아미노산의 합성 복잡한 경로를 통해 합성됨 5~16단계의 합성 경로 존재

28
2) 비필수 아미노산 (non-essential amino acid)
몸 안에서 합성되어 생기기 때문에, 따로 섭취해 줄 필요가 없는 아미노산 필수 아미노산 외의 아미노산 종류 글리신, 알라닌, 세린, 아스파라긴산, 글루탐산, 프롤린, 옥시프롤린, 아르기닌, 시스틴, 히스티딘, 티로신 등

29
비필수 아미노산의 합성 간단한 반응에 의해 합성됨 예외 Ex) 알라닌, 아스팔트산 알라닌→피루브산
아스팔트산→ 옥살로아세트산에서 한 단계를 거침으로써 합성됨 예외 아르기닌: 신생합성 경로 10단계 티로신 한 단계로 합성되기 때문에 비필수 아미노산으로 분류 처음부터 새로 합성되기 위해서 10단계 필요 페닐알라닌이 부족할 경우 필수 아미노산으로 분류

30
8종: 발린, 루신, 아이소루신, 메티오닌, 트레오닌, 리신,
아미노산(Amino acid) 필수 아미노산 Essential Amino Acid, EAA 비필수 아미노산 Non-essential amino acid 정의 생체 내에서 합성되지 않거나 합성이 어렵기 때문에 음식물로 섭취해야 하는 아미노산 몸 안에서 합성되어 생기기 때문에, 따로 섭취해 줄 필요가 없는 아미노산 종류 성인:8종, 영유아: 9종 8종: 발린, 루신, 아이소루신, 메티오닌, 트레오닌, 리신, 페닐알라닌,트립토판 9종: 영유아-히스티딘 글리신, 알라닌, 세린, 아스파라긴산, 글루탐산, 프롤린, 옥시프롤린, 아르기닌, 시스틴, 히스티딘, 티로신 등 합성 복잡한 경로를 통해 합성됨 합성 경로 단계: 5~16단계 간단한 반응에 의해 합성됨 합성 경로 단계: 1~5단계, 10단계 예외 티로신 한 단계로 합성되기 때문에 비필수 아미노산으로 분류 처음부터 새로 합성되기 위해서 10단계 필요 페닐알라닌이 부족할 경우 필수 아미노산으로 분류
 필수 아미노산. Essential Amino Acid, EAA. 비필수 아미노산. Non-essential. amino acid. 정의. 생체 내에서 합성되지 않거나. 합성이 어렵기 때문에. 음식물로 섭취해야 하는. 아미노산. 몸 안에서 합성되어 생기기. 때문에, 따로 섭취해 줄. 필요가 없는 아미노산. 종류. 성인:8종, 영유아: 9종. 8종: 발린, 루신, 아이소루신, 메티오닌, 트레오닌, 리신, 페닐알라닌,트립토판. 9종: 영유아-히스티딘. 글리신, 알라닌, 세린, 아스파라긴산, 글루탐산, 프롤린, 옥시프롤린, 아르기닌, 시스틴, 히스티딘, 티로신 등. 합성. 복잡한 경로를 통해 합성됨. 합성 경로 단계: 5~16단계. 간단한 반응에 의해 합성됨. 합성 경로 단계: 1~5단계, 10단계. 예외. 티로신. 한 단계로 합성되기 때문에 비필수 아미노산으로 분류. 처음부터 새로 합성되기 위해서 10단계 필요. 페닐알라닌이 부족할 경우 필수 아미노산으로 분류.")
31
2. 몇 가지 아미노산은 단순한 아미노기 전이반응에 의해서 만들어질 수 있다.
2. 몇 가지 아미노산은 단순한 아미노기 전이반응에 의해서 만들어질 수 있다. * α-케토산 ① α-케토글루타르산 ② 옥살로아세트산 ③ 피루브산 아미노기 전이반응 2종의 분자 사이에서 한 쪽에서 다른 쪽으로 아미노기가 이동하는 반응 아미노산의 amino 기를 제거하여 α-케토산 (α-keto acid) 수용체에 전달하는 것 하나의 아미노산으로부터 하나의 α-케토산으로의 아미노기의 전달에 의해 새로운 아미노산이 형성됨 비타민 B6 유도체인 피리독살인산을 보효소로 함 아미노기 이전효소로 불리는 효소에 의해서 촉매됨
 수용체에 전달하는 것. 하나의 아미노산으로부터 하나의. α-케토산으로의 아미노기의 전달에 의해. 새로운 아미노산이 형성됨. 비타민 B6 유도체인 피리독살인산을 보효소로 함. 아미노기 이전효소로 불리는 효소에 의해서 촉매됨.")
32
아미노기 전이반응 (Transamination reaction)
* 보충 설명 α-케토글루타르산 → → 글루탐산 환원 글루탐산의 아미노기 : 다른 α-케토글루타르산으로 이동 아미노기 결합 - 아스팔트산→옥살로아세트산 - 알라닌→피루브산 아미노기 전이반응 - 피리독살 인산 의존적 아미노기 전이효소에 의해 일어남 - 대부분의 아미노산 합성과정에 관여 옥살로아세트산 + 글루탐산 아스팔트산 + α-케토글루타르산 알라닌 + α-케토글루타르산 피루브산 + 글루탐산

33
아미노기 전이효소 (Transaminase, aminotransferase)
아미노산의 아미노기를 제거하여 다른 분자로 옮겨주는 반응을 촉매 하는 효소 아미노산과 α-케토산 사이에서 아미노기를 제거하여 다른 분자로 옮기는 생화학적인 반응을 촉매 하는 효소 보결분자단으로 피리독신(vitamin B6)에서 형성되는 피리독살 인산(pyridoxal phosphate, PLP)을 지니고 있음 * 보결분자단 복합단백질의 단백질에 결합되어 있는 비단백질성 분자 또는 분자단
에서 형성되는 피리독살 인산(pyridoxal phosphate, PLP)을 지니고 있음. * 보결분자단. 복합단백질의 단백질에 결합되어 있는. 비단백질성 분자 또는 분자단.")
34
피리독신 (vitamin B6), 피리독살 인산(PLP), 피리독사민 인산(PMP)
* 보충 설명 1. 피리독신(pyridoxine, vitamin B6 ) - 수용성비타민의 일종 - 피리독살 인산(PLP)과 피리독사민 인산(PMP), - 아미노기 전이효소를 포함 - 다수의 보조인자 2. 피리독살 인산(pyridoxal phosphate, PLP) - 피리독살이 피리독살인산화효소에 의해 활성형이 된 것 - 피리독신에서 만들어지는 조효소의 하나 - 아미노산의 아미노기 전달 반응, 카르복시 이탈, 축합 따위를 촉매하는 여러 효소의 조효소 - 피리독사민 인산(PMP)을 형성 3. 피리독사민 인산(pyridoxamine phosphate, PMP) - 아미노기외 결합하여 다양한 효소의 보조인자로 존재 - 피리독사민인산산화효소에 의해서 산화되어 피리독살5‘-인산으로 변환된다
 - 수용성비타민의 일종. - 피리독살 인산(PLP)과 피리독사민 인산(PMP), - 아미노기 전이효소를 포함. - 다수의 보조인자. 2. 피리독살 인산(pyridoxal phosphate, PLP) - 피리독살이 피리독살인산화효소에 의해 활성형이 된 것. - 피리독신에서 만들어지는 조효소의 하나. - 아미노산의 아미노기 전달 반응, 카르복시 이탈, 축합 따위를 촉매하는 여러 효소의 조효소. - 피리독사민 인산(PMP)을 형성. 3. 피리독사민 인산(pyridoxamine phosphate, PMP) - 아미노기외 결합하여 다양한 효소의 보조인자로 존재. - 피리독사민인산산화효소에 의해서 산화되어. 피리독살5‘-인산으로 변환된다.")
35
3. 세린, 시스테인, 글리신은 3-포스포글리세르산으로부터 합성된다
3. 세린, 시스테인, 글리신은 3-포스포글리세르산으로부터 합성된다 메티오닌아데노실전달효소에 의해 ATP와 메티오닌에서 합성 단백질이나 핵산이 메틸화할 때의 메틸기 공급체

36
H+ + NADH P O COO¯ 2¯ = P O H COO¯ OH 2¯ NAD+ 3-포스포글리세르산 3-포스포히드록시피루브산

37
P O COO¯ H 2¯ +H₃N P O COO¯ 2¯ = 글루탐산 α-케토글루타르산 3-포스포세린 3-포스포히드록시피루브산

38
P O COO¯ H 2¯ +H₃N H₂O Pi COO¯ H +H₃N OH 세린 3-포스포세린

39
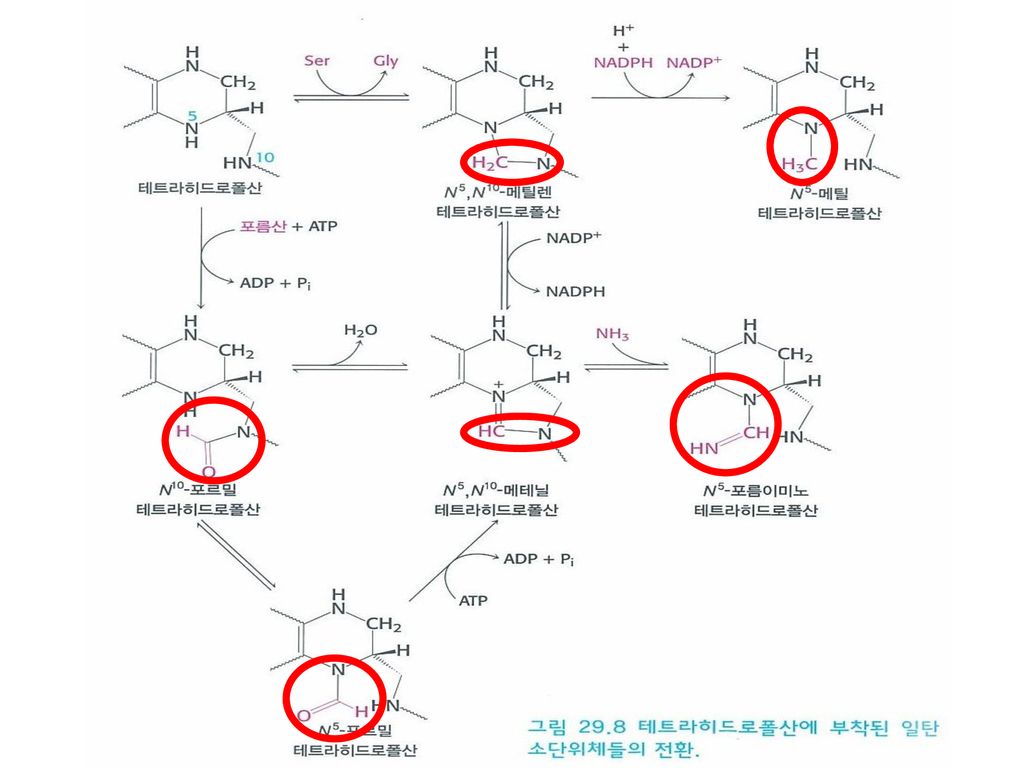
4. 테트라히드로폴산은 활성화된 하나의 탄소 단위를 운반한다
4. 테트라히드로폴산은 활성화된 하나의 탄소 단위를 운반한다 테트라히드로폴산 여러 아미노산들과 누클레오티드들의 합성에 필수적인 보조효소 활성된 일탄소 단위체의 운반체의 기능 엽산(비타민 B9)에서 형성됨 DNA복제과정과 세포의 성장에서 중요한 기능을 함 구성 테리딘(Pteridine) P-아미노벤조산(p-aminobenzoate) 글루탐산 잔기
에서 형성됨. DNA복제과정과 세포의 성장에서 중요한. 기능을 함. 구성. 테리딘(Pteridine) P-아미노벤조산(p-aminobenzoate) 글루탐산 잔기.")
40
테트라히드로폴산 구조식 ① 테리딘 ② P-아미노벤조산 ③ 글루탐산

41
테트라히드로폴산에 의해 운반되는 일탄소 단위체들
원자단 산화상태 구조식 이름 가장 많이 환원된 것 (=메탄올) -CH3 메틸 중간 정도로 환원 된 것 (=포름알데히드) -CH2- 메틸렌 가장 많이 산화 된 것 -CHO 포르밀 -CHNH 포름이미노 -CH= 메테닐
 -CH3. 메틸. 중간 정도로 환원 된 것. (=포름알데히드) -CH2- 메틸렌. 가장 많이 산화 된 것. -CHO. 포르밀. -CHNH. 포름이미노. -CH= 메테닐.")
43
5. S-아데노실메티오닌은 메틸기의 주요 증여자이다
(S-AdenosylcholineMethionine, SAM) ATP의 아데노실(adenosyl)기가 메티오닌 (methionine)의 황 원자에 결합하여 합성됨 활성화된 메틸기 제공자 포스파티딜에탄올아민(phosphatidylethanolamine)으로부터 포스파티콜린(phosphatidylcholine)을 합성하는 과정에 관여 에틸렌(ethylene)의 전구체 * 에틸렌(Ethylene) - 식물의 기체성 호르몬 - 열매를 익게 만드는 역할을 함 - 열매의 부패에 직접적으로 관여
 ATP의 아데노실(adenosyl)기가 메티오닌. (methionine)의 황 원자에 결합하여 합성됨. 활성화된 메틸기 제공자. 포스파티딜에탄올아민(phosphatidylethanolamine)으로부터. 포스파티콜린(phosphatidylcholine)을. 합성하는 과정에 관여. 에틸렌(ethylene)의 전구체. * 에틸렌(Ethylene) - 식물의 기체성 호르몬. - 열매를 익게 만드는 역할을 함. - 열매의 부패에 직접적으로 관여.")
44
S-아데노실메티오닌의 합성 * 메틸기 - 반응성 : 메티오닌의 메틸기 > N5-메틸렌테트라히드로폴산
- 주변의 황 원자의 양전하에 의해 활성화 됨 - 반응성 : 메티오닌의 메틸기 > N5-메틸렌테트라히드로폴산

45
S-아데노실메티오닌의 합성 * S-아데노실호모시스테인 - S-아데노실메티오닌의 메틸기가 수용체로 운반되어 형성 됨
메틸기가 수용체로 운반되어 형성 됨 - 호모시스테인과 아데노신으로 가수분해 됨 S-아데노실메티오닌의 합성

46
활성화된 메틸 회로 * 메티오닌의 메틸기 - S-아데노실메티오닌의 형성에 의해서 활성화 됨 * S-아데노실메티오닌
- 높은 메틸기 운반능력을 지님 → 다양한 종류의 수용체로 메틸기의 이동을 가능하게 만들어줌 * 메틸기(-CH₃) 호모시스테인이 메티오닌으로 전환되는 과정에서 회로에 참여 아데노실기의 결합에 의해 매우 반응적으로 변화함
 호모시스테인이 메티오닌으로. 전환되는 과정에서 회로에 참여. 아데노실기의 결합에 의해. 매우 반응적으로 변화함.")
47
29.3 되먹임 억제는 아미노산의 생합성을 조절한다 되먹임 억제(Feedback inhibition) 아미노산의 합성속도 결정
29.3 되먹임 억제는 아미노산의 생합성을 조절한다 되먹임 억제(Feedback inhibition) 여러 단계로 이루어진 효소계에서 최종단계의 생산물이 처음 단계의 효소를 억제하는 것 반응이 어느 정도 진행되면 정지하고 생성물은 일정한 농도를 유지함 각종 아미노산이나 누클레오티드의 합성계에서 일어남 아미노산의 합성속도 결정 ① 생합성 효소들의 양 ② 효소들의 활성정도
 여러 단계로 이루어진 효소계에서 최종단계의. 생산물이 처음 단계의 효소를 억제하는 것. 반응이 어느 정도 진행되면 정지하고 생성물은. 일정한 농도를 유지함. 각종 아미노산이나 누클레오티드의 합성계에서. 일어남. 아미노산의 합성속도 결정. ① 생합성 효소들의 양. ② 효소들의 활성정도.")
48
3-포스포글리세르산탈수소효소의 구조

49
1. 비가역적인 개입 단계는 일반적인 조절 단계이다
1. 비가역적인 개입 단계는 일반적인 조절 단계이다 A B D C E Z Z로 억제된다 생체 단위체와 물질대사 에너지의 보존에 필수적 * 개입단계(Committed step) = 비가역적 반응 → 물질대사 경로들에서 중요한 조절단계 - 대사경로의 최종 생성물 - 개입단계를 촉매 하는 효소억제
 = 비가역적 반응. → 물질대사 경로들에서. 중요한 조절단계. - 대사경로의 최종 생성물. - 개입단계를 촉매 하는. 효소억제.")
50
* 개입단계(Committed step)
대사경로 개입단계의 예시 세린 A B D C E Z Z로 억제된다 * 개입단계(Committed step) : 3-포스포글리세르산의 산화과정
 : 3-포스포글리세르산의 산화과정.")
51
* 세린+조절 도메인= 효소의 Vmax 값이 낮아짐
대사경로 개입단계의 예시 * 세린+조절 도메인= 효소의 Vmax 값이 낮아짐 비활성 상태→ 효소의 활성 억제 → 3-포스포글리세르산의 낭비를 방지 세린 3-포스포글리세르산 탈수소효소 - 대장균에서 4개의 동일한 구성단위체에 의해 사합체를 형성하고 있음 * 구성단위체 : 촉매 도메인과 세린 결합 조절 도메인을 → 조절 도메인 갖고 있음

52
2. 분지경로는 복잡하고 정교한 조절을 요구한다 되먹임 억제(피드백 억제)와 활성 아미노산-발린, 루신, 이소루신의 생합성
2. 분지경로는 복잡하고 정교한 조절을 요구한다 되먹임 억제(피드백 억제)와 활성 아미노산-발린, 루신, 이소루신의 생합성 과정 히드록시에틸 티아민 피로인산(TPP) - 3가지 아미노산들의 합성경로 시작물질 - 이소루신 합성경로의 시작단계에서 α-케토부틸산(ketobutyrare)과 반응함 - 발린과 루신 합성경로의 시작단계에서 피루브산과 반응 → α-케토부틸산과 피루브산의 농도 : 발린, 루신, 이소루신의 합성농도 결정
와 활성. 아미노산-발린, 루신, 이소루신의 생합성. 과정. 히드록시에틸 티아민 피로인산(TPP) - 3가지 아미노산들의 합성경로 시작물질. - 이소루신 합성경로의 시작단계에서. α-케토부틸산(ketobutyrare)과 반응함. - 발린과 루신 합성경로의 시작단계에서. 피루브산과 반응. → α-케토부틸산과 피루브산의 농도. : 발린, 루신, 이소루신의 합성농도 결정.")
53
트레오닌 탈아미노화효소의 조절 트레오닌 탈아미노화효소 α-케토부틸산 히드록시에틸-TTP 활성 억제 피루브산 루신 발린 이소루신
* 트레오닌 탈아미노화효소 - α-케토부틸산의 형성을 촉매 - 다른자리입체성으로 이소루신에 의해 억제 - 발린에 의해 다른자리입체성으로 활성화 됨 - 자신이 촉매하는 대사경로의 최종 생성물에 의해 억제 - 경쟁적인 대사경로의 최종 생성물에 의해 활성화 트레오닌 탈아미노화효소 α-케토부틸산 히드록시에틸-TTP 피루브산 활성 억제 루신 발린 이소루신

54
2) 효소의 다중도 (Enzyme multiplicity)
개입단계는 다른 조절성질을 갖은 두 개 또는 그 이상의 효소에 의해 촉매 됨 Ex) 트레오닌, 메티오닌, 리신의 생합성 과정 개입단계: 아스팔트산의 인산화 과정
 트레오닌, 메티오닌, 리신의 생합성 과정. 개입단계: 아스팔트산의 인산화 과정.")
55
효소의 다양성 X에 의해 억제 A X Y 효소 1 효소 2 Y에 의해 억제

56
3) 누적 되먹임 억제 (Cumulative feedback inhibition)
공통된 단계 : 부분적으로 각각의 최종 생성물에 독립적으로 억제되는 것 Ex) 대장균에서 글루타민 합성효소의 조절 글루타민=글루탐산+암모늄 이온+ATP 글루타민 합성효소 - 50kd의 동일한 12개의 구성단위체로 이루어짐 - 질소의 이동을 조절 - 박테리아의 물질대사 조절에서 중요한 역할 - 알라닌, 글리신, 글루타민 물질대사의 최종 생성물에 의해 누적적으로 억제 → 누적 억제: 효소의 활성을 억제
 대장균에서 글루타민 합성효소의 조절. 글루타민=글루탐산+암모늄 이온+ATP. 글루타민 합성효소. - 50kd의 동일한 12개의 구성단위체로 이루어짐. - 질소의 이동을 조절. - 박테리아의 물질대사 조절에서 중요한 역할. - 알라닌, 글리신, 글루타민 물질대사의. 최종 생성물에 의해 누적적으로 억제. → 누적 억제: 효소의 활성을 억제.")
57
Chapter 30 누클레오티드 대사

58
목 차 2. Chapter 30 누클레오티드 대사 30.2 피리미딘 고리는 조립되어 리보오스 당에 붙는다.
목 차 2. Chapter 30 누클레오티드 대사 30.1 누클레오티드 생합성과 명명법에 대한 개요 30.2 피리미딘 고리는 조립되어 리보오스 당에 붙는다. 30.3 퓨린 고리는 리보오스 인산 위에 조절된다. 30.4 리보누클레오티드들은 데옥시리보누클레오티드들로 환원된다. 30.5 누클레오티드 생합성은 되먹임 억제 (feedback inhibition) 에 의해 조절된다. 30.6 누클레오티드 대사의 혼란은 병리적 상태를 야기할 수있다.
 에 의해 조절된다 누클레오티드 대사의 혼란은 병리적 상태를. 야기할 수있다.")
59
누클레오시드와 티드의 정의 RNA DNA 염기 1. 뉴클레오시드 : 5탄당 1분자 + 염기 1분자 아데닌[A]
리보 누클레오 시드 리보누클레오티드 5-일인산 데옥시 누클레오 시드 티드 아데닌[A] 아데노신 아데닐산 [AMP] [dAMP] 구아닌[G] 구아노신 구아닐산 [GMP] [dGMP] 우라실[U] 우리딘 우리딜산 [UMP] 티민[T] 티미딘 티미디산 [TMP] 시토신[C] 시티신 시티딜산 [CMP] 데옥시 시티딘 [dCMP] 누클레오시드와 티드의 정의 1. 뉴클레오시드 : 5탄당 1분자 + 염기 1분자 - 5탄당이 리보당일 때는 RNA이고 5탄당이 데옥시리보당일 때는 DNA를 만들 수 있음 염기 종류 - DNA의 구성: A[아데닌], T[티민], G[구아닌], C[시토신] - RNA의 구성: A[아데닌], U[우라실], G[구아닌], C[시토신] 단일환상구조로 된 피리미딘계 : 시토신, 우라실, 티민 이중환상구조로 된 퓨린계 : 아데닌, 구아닌 2.뉴클레오티드 : 뉴클레오시드에 인산이 에스테르 결합된 화합물 표.30.1 염기 누클레오시드 그리고 누클레오티드의 명명법
![누클레오시드와 티드의 정의 RNA DNA 염기 1. 뉴클레오시드 : 5탄당 1분자 + 염기 1분자 아데닌[A]](http://slidesplayer.org/slide/14985287/91/images/59/%EB%88%84%ED%81%B4%EB%A0%88%EC%98%A4%EC%8B%9C%EB%93%9C%EC%99%80+%ED%8B%B0%EB%93%9C%EC%9D%98+%EC%A0%95%EC%9D%98+RNA+DNA+%EC%97%BC%EA%B8%B0+1.+%EB%89%B4%ED%81%B4%EB%A0%88%EC%98%A4%EC%8B%9C%EB%93%9C+%3A+5%ED%83%84%EB%8B%B9+1%EB%B6%84%EC%9E%90+%2B+%EC%97%BC%EA%B8%B0+1%EB%B6%84%EC%9E%90+%EC%95%84%EB%8D%B0%EB%8B%8C%5BA%5D.jpg "리보. 누클레오 시드. 리보누클레오티드. 5-일인산. 데옥시. 누클레오. 시드. 티드. 아데닌[A] 아데노신. 아데닐산. [AMP] [dAMP] 구아닌[G] 구아노신. 구아닐산. [GMP] [dGMP] 우라실[U] 우리딘. 우리딜산. [UMP] 티민[T] 티미딘. 티미디산. [TMP] 시토신[C] 시티신. 시티딜산. [CMP] 데옥시 시티딘. [dCMP] 누클레오시드와 티드의 정의. 1. 뉴클레오시드 : 5탄당 1분자 + 염기 1분자. - 5탄당이 리보당일 때는 RNA이고 5탄당이 데옥시리보당일 때는. DNA를 만들 수 있음. 염기 종류. - DNA의 구성: A[아데닌], T[티민], G[구아닌], C[시토신] - RNA의 구성: A[아데닌], U[우라실], G[구아닌], C[시토신] 단일환상구조로 된 피리미딘계 : 시토신, 우라실, 티민. 이중환상구조로 된 퓨린계 : 아데닌, 구아닌. 2.뉴클레오티드 : 뉴클레오시드에 인산이 에스테르 결합된. 화합물. 표.30.1 염기 누클레오시드 그리고 누클레오티드의 명명법.")
60
활성화된 리보오스(PRPP) + 아미노산들 + ATP + Co2 + …
30.1 누클레오티드 생합성 신생 경로 활성화된 리보오스(PRPP) + 아미노산들 + ATP + Co2 + … 회수 경로 활성화된 리보오스(PRPP) + 염기 누클레오티드 누클레오티드
 + 아미노산들 + ATP + Co2 + … 회수 경로. 활성화된 리보오스(PRPP) + 염기. 누클레오티드. 누클레오티드.")
61
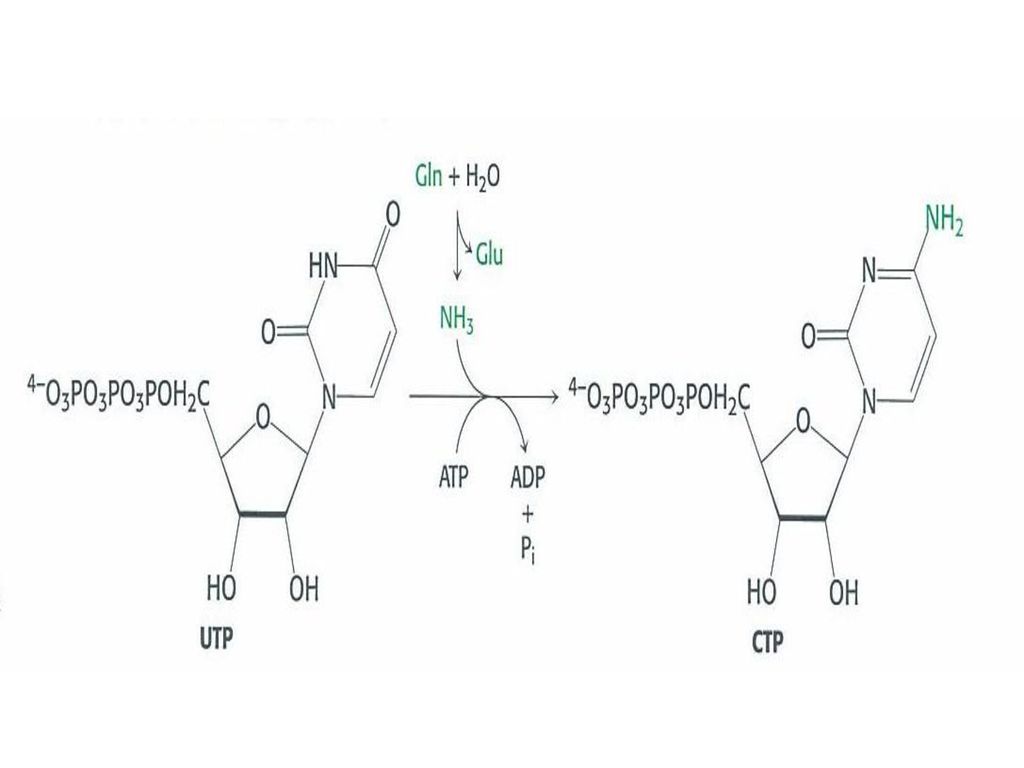
30.2 피리미딘 고리는 조립되어 리보오스 당에 붙는다.
30.2 피리미딘 고리는 조립되어 리보오스 당에 붙는다. 피리미딘의 신생합성은 중탄산염과 암모니아가 2개의 ATP를 소모해서 카르모바일 인산을 만들고 아스팔트산과 만나서 피라미딘 고리를 만들며, 피리미딘 고리가 PRPP와 결합하여 우리딜산을 만들고 UMD는UTP가 되서 [UTP], [CTP], [dCTP], [TTP]

62
1. 키나아제는 누클레오시드 일인산을 누클레오시드 삼인산으로 변환한다
1. 키나아제는 누클레오시드 일인산을 누클레오시드 삼인산으로 변환한다 UMP + ATP ⇄ UDP + ADP ↑ XDP + YTP ⇄ XTP + YDP UMP 키나아제 효소 누클레오시드 이인산 키나아제 효소에 의해 상호호환

64
퓨린 고리는 처음부터 당이 붙고 9개 과정을 거친 후 IMP가 되서 AMP와 GMP가 된다.
30.3 퓨린 고리는 리보오스 인산 위에 조립된다 퓨린 고리는 처음부터 당이 붙고 9개 과정을 거친 후 IMP가 되서 AMP와 GMP가 된다.

65
1. AMP와 GMP는 IMP로부터 형성된다 ¯OOC 아데닐로석신산 이노신산 잔틸산 GDP Pi GTP + Asp COO¯ O
N P-리보오스 HN ¯OOC COO¯ H 아데닐로석신산 N NH O P-리보오스 이노신산 N O P-리보오스 잔틸산 H NH NAD + H2O NADH H

66
¯OOC ¯OOC 푸마르산 P-리보오스 아데닐로석신산 아데닐산(AMP) NH2 N COO¯ COO¯ H H HN N
 NH2 N COO¯ COO¯ H H HN N")
67
P-리보오스 잔틸산 구아닐산(GMP) H3N+ H2O H2O+ Gln ATP AMP + PPi N O NH2 O N NH
 H3N+ H2O H2O+ Gln ATP AMP + PPi N O NH2 O N NH")
68
2. 염기는 회수경로에 의해 재이용될 수 있다 아데닌 + PRPP → AMP + PPi
구아닌 + PRPP → GMP + PPi 히포잔틴 + PRPP → 이노신산 + PPi

69
접두어 용어 설명 Ribo- : Ribose의 뜻 Deoxy- : 탈산소의 뜻 발음→ 데옥시 또는 디옥시
Nucle- (자음 앞에서는 Nucleo-) : 핵, 핵산의 뜻 발음→ 누클 또는 뉴클
 : 핵, 핵산의 뜻. 발음→ 누클 또는 뉴클.")
70
30.4 리보누클레오티드들은 데옥시리보누클레오티드들로 환원된다
30.4 리보누클레오티드들은 데옥시리보누클레오티드들로 환원된다 1) 리보누클레오티드(Ribonucleotide) Ribose에 purine기 또는 pyrimidine기가 → 다른 리보누클레오시드 및 누클레오티드, NAD+와 보조효소 A 같은 보조인자의 탄수화물 부분을 구성하는 5탄당 연결되어 이루어진 물질 당 부분이 D-리보오스인 것 RNA의 가수분해에 의해 얻어짐 Ribose를 함유한 누클레오티드(Nucleotide)
 리보누클레오티드(Ribonucleotide) Ribose에 purine기 또는 pyrimidine기가. → 다른 리보누클레오시드 및 누클레오티드, NAD+와 보조효소 A 같은 보조인자의 탄수화물 부분을. 구성하는 5탄당. 연결되어 이루어진 물질. 당 부분이 D-리보오스인 것. RNA의 가수분해에 의해 얻어짐. Ribose를 함유한 누클레오티드(Nucleotide)")
71
2) 누클레오시드(Nucleoside) 염기와 오탄당만 있고, 인산이 없는 물질 Nucleoside= 염기+오탄당
= Ribose + Base - Purine: Adenine, Guanine - Pyrinidine: Uracil, Cytosine, Thymine

72
* Nucleoside와 Nucleotide의 차이
핵산 구성성분의 최소단위 염기, 당, 인산의 3가지 요소로 구성된 화학적 단량체 DNA나 RNA 같은 핵산을 이루는 단위체 (핵산의 구성성분) Nucleotide= Nucleoside + Phosphate = 염기 + 오탄당 + 인산 염기 - 퓨린 계열: 아데닌(Adenine, A), 구아닌(Guanine, G) - 피리미딘 계열: 시토신(Cytosine, C), 티민(Thymine, T), 우라실(Uracil, U) 오탄당 - 리보오스(Ribose) - 디옥시리보오스(Deoxyribose) 인산 * Nucleoside와 Nucleotide의 차이 - 인산의 유뮤 - 인산이 있음→ Nucleotide Nucleotide= Nucleoside + Phosphate = 염기 + 오탄당 + 인산 - 인산이 없음→ Nucleoside Nucleoside= 염기+오탄당 = Ribose + Base
 Nucleotide= Nucleoside + Phosphate. = 염기 + 오탄당 + 인산. 염기. - 퓨린 계열: 아데닌(Adenine, A), 구아닌(Guanine, G) - 피리미딘 계열: 시토신(Cytosine, C), 티민(Thymine, T), 우라실(Uracil, U) 오탄당 - 리보오스(Ribose) - 디옥시리보오스(Deoxyribose) 인산. * Nucleoside와 Nucleotide의 차이. - 인산의 유뮤. - 인산이 있음→ Nucleotide. Nucleotide= Nucleoside + Phosphate. = 염기 + 오탄당 + 인산. - 인산이 없음→ Nucleoside. Nucleoside= 염기+오탄당. = Ribose + Base.")
73
누클레오티드(Nucleotide)의 종류와 약기
DNA 디옥시아데닐산 dAMP 디옥시구아닐산 dGMP 디옥시시티딜산 dCMP (디옥시)티미딜산 TMP RNA 아데닐산 AMP 구아닐산 GMP 시티딜산 CMP 우리딜산 UMP
티미딜산. TMP. RNA. 아데닐산. AMP. 구아닐산. GMP. 시티딜산. CMP. 우리딜산. UMP.")
74
4) 데옥시리보누클레오티드 (Deoxyribonucleotide)
데옥시리보오스를 포함한 DNA 구성요소 → deoxyribose: 데옥시누클레오티드의 탄수화물 부분을 구성하는 5탄소의 단당류 DNA의 전구체 리보누클레오시드 이인산의 환원에 의해 형성됨 당 부분이 D-2’-데옥시리보오스인 것 DNA의 효소분해를 통해 얻어짐

75
5) 환원(Reduction) 산화된 것을 원래의 상태로 돌리는 것 산화의 역 과정 어떤 물질이 산소를 잃어버리거나 수소를
얻는 것 원소와 이온이 전자를 받아들여 +원자가를 빼거나 –원자가를 늘리는 것

76
1. 데옥시리보누클레오티드의 형성 ADP GDP CDP UDP 리보누클레오티드 환원효소 데옥시리보누클레오티드2인산 dADP
아데노신2인산 구아노신2인산 시티딘2인산 우리딘2인산 ADP GDP CDP UDP 리보누클레오티드 환원효소 데옥시리보누클레오티드2인산 dADP dGDP dCDP dUDP dATP dGTP dCTP TTP 데옥시아데노신 3인산 데옥시구아노신 데옥시티딘 데옥시티미딘 리보누클레오티드 환원효소의 생성물들 추가적인 과정을 통해 dNTP를 얻는다

77
용어 설명 데옥시리보누클레오시드 (deoxyribonucleoside) DNA를 구성하고 있는 Nucleoside
데옥시리보오스와 퓨린 또는 피리미딘 염기로 이루어짐 4종류 데옥시아데노신 데옥시구아닌(구아노신) 데옥시시티딘 데옥시티미딘
 데옥시시티딘. 데옥시티미딘.")
78
용어 설명 2) 리보누클레오티드 환원효소 (Ribonucleotide reductase) 모든 생물에 필수적인 존재
DNA 생합성에 있어 중심적 역할을 담당하는 효소 데옥시체(dNTP)로 만드는 효소 → 리보누클레오시드 3인산환원효소
로 만드는 효소. → 리보누클레오시드 3인산환원효소.")
79
2. 데옥시리보누클레오티드의 합성 (1) 리보누클레오티드 이인산+NADPH+H+ 리보누클레오티드 환원효소 데옥시리보누클레오티드 이인산+NADP++H₂O * 티오레독신(Thioredoxin, TRX) NADPH와 환원효소를 연결하는 환원력의 운반체 리보누클레오티드 환원효소가 리보누클레오티드를 환원할 때의 Proton 공급체 →수소원자에서 전자가 상실되어 형성되는 수소 양이온

80
데옥시리보누클레오티드의 합성 (2) SH 리보누클레오시드 이인산+R SH S S 리보누클레오시드 환원효소
* 이황화 결합 2개의 설프히드릴기(-SH)의 산화에 의해 형성되는 결합 (S-S결합) → 리보누클레오티드 환원효소의 촉매작용에 의해 이황화물로 산화됨 * 설프히드릴기 티올기(-SH)의 별명 단백질의 시스테인 잔기의 SH기를 나타내는 경우에 자주 사용됨 S S
의. 산화에 의해 형성되는 결합. (S-S결합) → 리보누클레오티드 환원효소의. 촉매작용에 의해 이황화물로. 산화됨. * 설프히드릴기. 티올기(-SH)의 별명. 단백질의 시스테인 잔기의. SH기를 나타내는 경우에. 자주 사용됨. S. S.")
81
데옥시리보누클레오티드의 합성 (3) SH FAD TR S FADH2 NADP 티오레독신 환원효소 ( TR )
* 전자의 흐름: NADPH → FAD FAD FADH2 TR SH S 티오레독신 환원효소 ( TR ) NADPH + H + NADP NADPH로부터 나온 전자로 인해 환원된 티오레독신이 재생성되는 반응을 촉진
 NADPH + H. + NADP. NADPH로부터 나온 전자로 인해 환원된. 티오레독신이 재생성되는 반응을 촉진.")
82
→ 리보누클레오티드환원효소→리보오스 단위체
* 이황화물 결합 2개의 SH가 산화되어 생성하는 –S-S 형태의 황 원자 간 결합 * 전자의 흐름: FAD→ 티오레독신의 이황화물 → 리보누클레오티드환원효소→리보오스 단위체 데옥시리보누클레오티드의 합성 (3) * 이황화물 -S-S-기를 포함하고 있는 유기화합물 T SH S 티오레독신 ( T ) SH PR S 리보누클레오티드 환원효소 리보오스 단위체 데옥시리보오스 산화
 * 이황화물. -S-S-기를 포함하고 있는 유기화합물. T. SH. S. 티오레독신. ( T ) SH. PR. S. 리보누클레오티드. 환원효소. 리보오스. 단위체. 데옥시리보오스. 산화.")
83
3. 티미딜산은 데옥시우리딜산의 메틸화에 의해서 형성된다
3. 티미딜산은 데옥시우리딜산의 메틸화에 의해서 형성된다 1) 티미딜산(Thymidylate, TMP) 데옥시리보누클레오티드의 하나 티민과 데옥시리보오스와 무기인산으로 구성된 누클레오티드 2) 데옥시우리딜산(Deoxyuridylate, dUMP) 우라실의 데옥시리보누클레오티드 티미딜산(TMP)으로 메틸화 됨 dTMP합성효소로 메틸화하여 dTMP가 됨 → dTDP, dTTP의 합성에 사용 티미딘 인산화효소의 작용으로 데옥시우리딘에서 생성됨 * 메틸화(Methylation) 유기화합물의 탄소, 질소, 산소, 황 원자 등과 결합한 수소원자를 메틸기 (-CH3)로 치환하는 반응
 티미딜산(Thymidylate, TMP) 데옥시리보누클레오티드의 하나. 티민과 데옥시리보오스와 무기인산으로. 구성된 누클레오티드. 2) 데옥시우리딜산(Deoxyuridylate, dUMP) 우라실의 데옥시리보누클레오티드. 티미딜산(TMP)으로 메틸화 됨. dTMP합성효소로 메틸화하여 dTMP가 됨. → dTDP, dTTP의 합성에 사용. 티미딘 인산화효소의 작용으로. 데옥시우리딘에서 생성됨. * 메틸화(Methylation) 유기화합물의 탄소, 질소, 산소, 황 원자 등과 결합한 수소원자를. 메틸기 (-CH3)로 치환하는 반응.")
84
우라실로부터 티미딜산을 생성하기 위한 단계 티미딜산 생성(합성)효소가 촉진 티미딜산 생성(합성)효소
우라실로부터 티미딜산을 생성하기 위한 단계 티미딜산 생성(합성)효소가 촉진 티미딜산 생성(합성)효소 (Thymidylate synthase) 데옥시우리딜산의 5위의 탄소를 메틸화한 티미딜산을 생성하는 반응을 촉매하는 효소 DNA에 특이적 염기인 티민을 공급하는 역할 DNA전구체 공급경로의 주요한 속도조절효소의 일종 N5, N10-메틸렌테트라히드로폴산 (N5, N10-methylenetetrahydrofolate)이 메틸기의 공급체가 됨
효소가 촉진. 티미딜산 생성(합성)효소. (Thymidylate synthase) 데옥시우리딜산의 5위의 탄소를 메틸화한. 티미딜산을 생성하는 반응을 촉매하는 효소. DNA에 특이적 염기인 티민을 공급하는 역할. DNA전구체 공급경로의 주요한 속도조절효소의 일종. N5, N10-메틸렌테트라히드로폴산. (N5, N10-methylenetetrahydrofolate)이 메틸기의. 공급체가 됨.")
85
용어 설명 1) 폴산(Folic acid, 엽산) 수용성 비타민 B군의 일종 엽산이라고도 함
5, 6, 7, 8-테트라히드로엽산의 형태로 생체 내의 포르밀기, 메테닐기, 메틸렌기, 메틸기, 포름이미노기 등 각종 C₁단위의 전달 반응에 관여하는 효소의 조효소가 됨 퓨린, 피리미딘의 대사와 글리신, 세린, 히스티딘, 메티오닌 등의 아미노산 대사, 단백질의 합성개시, 메탄생성 등에 관여

86
용어 설명 2) N5, N10-메틸렌테트라히드로폴산(엽산) (N5, N10-methylenetetrahydrofolate)
데옥시우리딜산(dUMP)이 티미딜산(TMP)으로 메틸화 되는 반응에서 메틸기의 공여체 메틸전달효소→메틸기 전달효소의 일종
이 티미딜산(TMP)으로 메틸화 되는 반응에서 메틸기의 공여체. 메틸전달효소→메틸기 전달효소의 일종.")
87
티미딜산의 형성과정 티미딜산을 생성하는 반응을 촉매하는 효소

88
* 테트라히드로폴산(Terahydrofolic acid)
테트라히드로폴산의 형성과정 * 테트라히드로폴산(Terahydrofolic acid) - 테트라히드로프로테일 글루탐산에 해당하는 엽산의 조효소형 - 엽산의 환원체 - 공기에 닿으면 쉽게 산화되는 물질 - 엽산의 NADPH에 의한 2단계 환원에 의해 생성됨 * 재생성 NADPH+디히드로폴산 환원효소 티미딜산의 합성으로 생성됨 디히드로폴산으로부터 재생성 됨
 - 테트라히드로프로테일. 글루탐산에 해당하는 엽산의. 조효소형. - 엽산의 환원체. - 공기에 닿으면 쉽게 산화되는. 물질. - 엽산의 NADPH에 의한 2단계. 환원에 의해 생성됨. * 재생성. NADPH+디히드로폴산 환원효소. 티미딜산의 합성으로 생성됨. 디히드로폴산으로부터 재생성 됨.")
89
30.5 누클레오티드 생합성은 되먹임 억제에 의해 조절된다
30.5 누클레오티드 생합성은 되먹임 억제에 의해 조절된다 되먹임 억제(Feedback infibition) 대사제어 기구의 하나 여러 단계로 이루어진 효소계에서 최종단계의 생산물이 처음 단계의 효소를 억제하는 것 반응이 어느 정도 진행되면 정지하고 생성물은 일정한 농도를 유지함 각종 아미노산이나 누클레오티드의 합성계에서 일어남
 대사제어 기구의 하나. 여러 단계로 이루어진 효소계에서 최종단계의 생산물이 처음 단계의 효소를 억제하는 것. 반응이 어느 정도 진행되면 정지하고 생성물은 일정한 농도를 유지함. 각종 아미노산이나 누클레오티드의 합성계에서 일어남.")
90
1. 피리미딘 생합성은 아스팔트산 카르바모일기 전이효소에 의해 조절된다.
1. 피리미딘 생합성은 아스팔트산 카르바모일기 전이효소에 의해 조절된다. 아스팔트산 카르바모일기 전이효소 (Aspartate transcarbamoylase, ATCase) 피리미딘 생합성을 조절하는 주요효소 억제: CTP→피리미딘 생합성의 마지막 생성물 자극: ATP
 피리미딘 생합성을 조절하는 주요효소. 억제: CTP→피리미딘 생합성의 마지막 생성물. 자극: ATP.")
91
→ + - 아스팔트산 + 카르바모일 인산 ATCase ATP 카르바모일아스팔트산 →→→UMP→UDP→UTP→CTP 자극
= + + = + - * 카르바모일 인산 아스팔트산과 반응하여 카르바모일아스팔트산을 생성함 → 자극 * ATP (아데노신3인산) 아스팔트산 카르바모일기 전이효소를 촉진 ATCase ATP * ATCase (아스팔트산 카르바모일기 전이효소) ATP에 의해 자극됨 개입 단계를 촉매하는 효소 CTP에 의해 억제됨 카르바모일아스팔트산 * CTP (시티딘3인산) 아스팔트산 카르바모일기 전이효소를 억제 →→→UMP→UDP→UTP→CTP 우리딜산 우리딘2인산 우리딘3인산 시티딘3인산
 아스팔트산 카르바모일기. 전이효소를 촉진. ATCase. ATP. * ATCase. (아스팔트산 카르바모일기. 전이효소) ATP에 의해 자극됨. 개입 단계를 촉매하는 효소. CTP에 의해 억제됨. 카르바모일아스팔트산. * CTP (시티딘3인산) 아스팔트산 카르바모일기. 전이효소를 억제. →→→UMP→UDP→UTP→CTP. 우리딜산 우리딘2인산 우리딘3인산 시티딘3인산.")
92
2. 퓨린 누클레오티드의 합성은 몇 가지 부위의 되먹임 억제에 의해 조절된다
1) 이노신산(Inosine MonoPhosphate, IMP) AMP(아데닌산)와 GMP(구아닐산) 모두의 전구체인 5-포스포리보실-1-피로인산(PRPP)과 히포잔틴의 반응에 의해 형성되는 퓨린 누클레오티드 GMP(구아닐산)의 형성 촉진 고기 엑기스에서 분리된 근육아데닐산의 탈아미노체 육즙의 단맛 성분, 조미료로서 사용함 퓨린누클레오티드의 합성 과정 중 만들어짐 (이노신산→아데닐산→ 구아닐산으로 변환됨)
 이노신산(Inosine MonoPhosphate, IMP) AMP(아데닌산)와 GMP(구아닐산) 모두의. 전구체인 5-포스포리보실-1-피로인산(PRPP)과. 히포잔틴의 반응에 의해 형성되는 퓨린. 누클레오티드. GMP(구아닐산)의 형성 촉진. 고기 엑기스에서 분리된 근육아데닐산의. 탈아미노체. 육즙의 단맛 성분, 조미료로서 사용함. 퓨린누클레오티드의 합성 과정 중 만들어짐. (이노신산→아데닐산→ 구아닐산으로 변환됨)")
93
(Adenosine MonoPhosphate, AMP)
2) 아데닐산 (Adenosine MonoPhosphate, AMP) 아데노신의 리보오스에 인산 1분자가 결합한 누클레오티드 RNA의 성분 구성: 인산기, 리보오스(당), 아데닌(핵염기) ADP, ATP를 만듦 각종 조효소의 성분 인산과 결합함으로써 고에너지를 갖는 ADP와 ATP로 되고, 역으로 이들의 분해로 AMP를 생성함
 아데닐산. (Adenosine MonoPhosphate, AMP) 아데노신의 리보오스에 인산 1분자가 결합한. 누클레오티드. RNA의 성분. 구성: 인산기, 리보오스(당), 아데닌(핵염기) ADP, ATP를 만듦. 각종 조효소의 성분. 인산과 결합함으로써 고에너지를 갖는 ADP와. ATP로 되고, 역으로 이들의 분해로 AMP를. 생성함.")
94
3) 구아닐산(Guanosin MonoPhosphate, GMP)
구아노신 일인산 누클레오티드의 하나 퓨린염기의 리보시드인 구아노신에 인산 1분자가 에스테르 결합한 것

95
퓨린 생합성의 조절 되먹임 억제에 의해 조절된다 * AMP(아데닐산) 이노신산이 * IMP(이노신산) 아데닐로석신산으로
전환되는 것을 억제 최종단계의 생산물 (최종산물) * IMP(이노신산) - AMP(아데닐산)와 GMP(구아닐산) 합성의 분기점 - GMP(구아닐산)의 형성 촉진 * AMP(아데닐산), GMP(구아닐산) 아미드기 전이효소를 억제 * 퓨린 누클레오티드의 합성은 몇 가지 부위의 되먹임 억제에 의해 조절된다 * 되먹임 억제 - 퓨린 생합성의 전체적인 속도 조절 - AMP(아데닐산)과 GMP(구아닐산) 사이의 균형 조절 5-포스포리보실-1피로인산 * 포스포리보실아민 Nucleoside+아민기 * GMP(구아닐산) 이노신산이 잔틸산으로 전환되는 것을 억제 되먹임 억제의 부위
 * IMP(이노신산) - AMP(아데닐산)와 GMP(구아닐산) 합성의 분기점. - GMP(구아닐산)의 형성 촉진. * AMP(아데닐산), GMP(구아닐산) 아미드기 전이효소를 억제. * 퓨린 누클레오티드의 합성은 몇 가지 부위의. 되먹임 억제에 의해 조절된다. * 되먹임 억제. - 퓨린 생합성의 전체적인 속도 조절. - AMP(아데닐산)과 GMP(구아닐산) 사이의 균형 조절. 5-포스포리보실-1피로인산. * 포스포리보실아민. Nucleoside+아민기. * GMP(구아닐산) 이노신산이 잔틸산으로. 전환되는 것을 억제. 되먹임 억제의. 부위.")
96
퓨린 생합성의 조절 ① 퓨린 누클레오티드 생합성의 개입단계 PRPP 포스포리보실아민 글루타민 포스포리보실 아미드기 전이효소
전환

97
② IMP(이노신산) AMP(아데닌산)와 GMP(구아닐산) 합성의 분기점 IMP(이노신산)으로부터 갈라져 나가는 반응
아데닐로석신산으로 전환되는 것을 억제 GMP(구아닐산) 이노신산이 GMP의 가장 가까운 전구체인 잔틸산으로 전환되는 것을 억제 IMP(이노신산)으로부터 갈라져 나가는 반응 → 되먹임 억제의 부위
 이노신산이 GMP의 가장 가까운 전구체인. 잔틸산으로 전환되는 것을 억제. IMP(이노신산)으로부터 갈라져. 나가는 반응. → 되먹임 억제의 부위.")
98
③ GTP(구아닐3인산) : AMP(아데닐산) 합성의 기질 상반되는 기질 사이의 관계 ATP(아데노신3인산)
: GMP(구아닐산) 합성의 기질 ∴ 아데닌과 구아닌 리보누클레오티드의 합성이 균형을 이루도록 함
 합성의 기질. ∴ 아데닌과 구아닌 리보누클레오티드의. 합성이 균형을 이루도록 함.")
99
3. 데옥시리보누클레오티드의 합성은 리보누클레오티드 환원효소의 조절에 의해 조정된다.
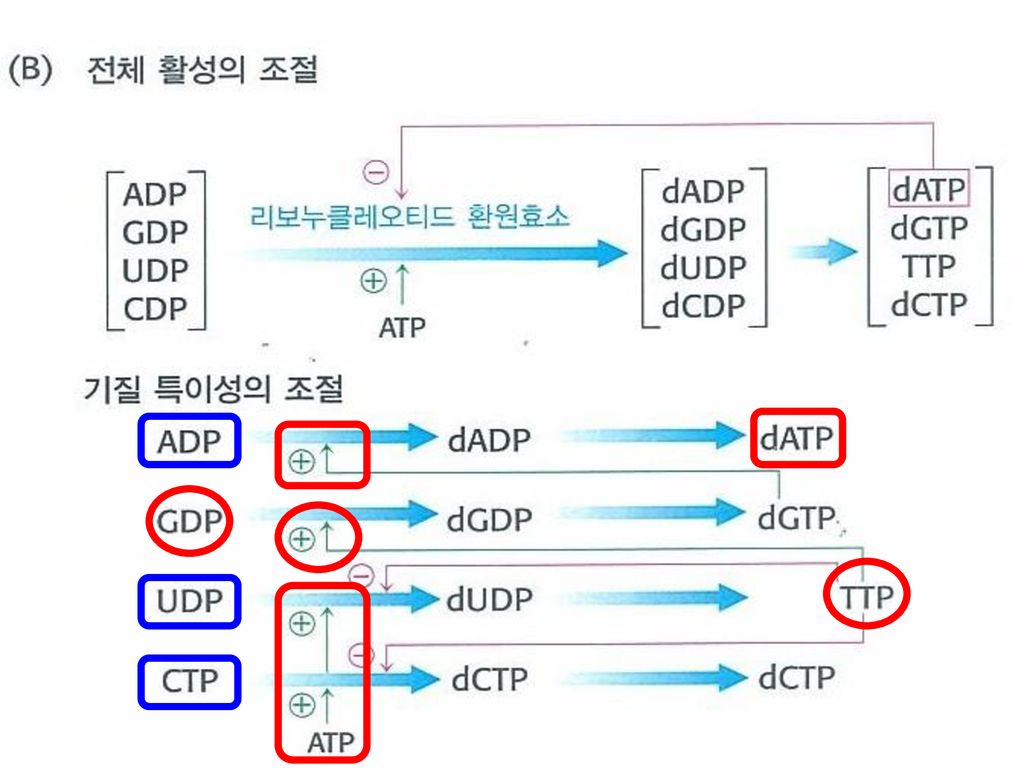
3. 데옥시리보누클레오티드의 합성은 리보누클레오티드 환원효소의 조절에 의해 조정된다. 리보누클레오티드 데옥시리보누클레오티드 환원 다른자리입체성 상호작용에 의해 조절됨

100
리보누클레오티드 환원효소 단일이량체효소 2개의 다른자리입체성 부위 ① 전체적인 반응성 조절 부위 ② 기질특이성 조절 부위
전체적인 촉매 활성도 dATP와의 결합에 의해 감소됨 → dATP: 데옥시리보누클레오티드가 풍부하다는 신호로 작용 ATP의 결합: 되먹임 억제를 뒤바꿈

101
리보누클레오티드 환원효소의 조절 전체적인 반응성 조절 기질 특이성 조절

102
기질 특이성을 조절하는 부위 + ① dATP 또는 ATP 결합 ② TTP(티미딘3인산)의 결합 ③ dGTP의 증가
UDP와 CDP같은 피리미딘 누클레오티드들의 환원이 증가 ② TTP(티미딘3인산)의 결합 GDP의 환원을 촉진 피리미딘 리보누클레오티드들이 더 환원되는 것을 억제 ③ dGTP의 증가 ATP가 dATP로 환원되도록 촉진
의 결합. GDP의 환원을 촉진. 피리미딘 리보누클레오티드들이. 더 환원되는 것을 억제. ③ dGTP의 증가. ATP가 dATP로 환원되도록 촉진.")
104
30.6 누클레오티드 대사의 혼란은 병리적 상태를 야기할 수 있다
누클레오티드 누클레오시드 누클레오시드 + 자유 염기 분해 누클레오티드 가수분해효소 가인산분해 리보오스 1-인산 또는 데옥시리보오스 1-인산 누클레오시드 가인산분해효소

105
퓨린의 이화작용(1) N H₂N N H₂N H₂O Pi H₂O NH4 + 리보오스5-P 리보오스 AMP N NH O 아데노신
* 이화작용(catabolism) - 생물이 체내에서 고분자유기물을 좀 더 간단한 저분자유기물이나 무기물로 분해하는 과정 - 생체내의 유기화합물이 분해하고 그 때 에너지를 유리하거나 열을 발생하거나 하는 현상 - 에너지를 얻기 위한 생체 반응 N H₂N N H₂N H₂O Pi H₂O 누클레오티드 가수분해효소 탈아미노화효소 아데노신 NH4 + 리보오스5-P 리보오스 AMP N NH O 아데노신 N NH H O 리보오스1-P Pi N 누클레오시드 인산화효소 리보오스 히포잔틴 이노신
 - 생물이 체내에서 고분자유기물을. 좀 더 간단한 저분자유기물이나 무기물로. 분해하는 과정. - 생체내의 유기화합물이 분해하고. 그 때 에너지를 유리하거나. 열을 발생하거나 하는 현상. - 에너지를 얻기 위한 생체 반응. N. H₂N. N. H₂N. H₂O. Pi. H₂O. 누클레오티드. 가수분해효소. 탈아미노화효소. 아데노신. NH4 + 리보오스5-P. 리보오스. AMP. N. NH. O. 아데노신. N. NH. H. O. 리보오스1-P. Pi. N. 누클레오시드. 인산화효소. 리보오스. 히포잔틴. 이노신.")
106
퓨린의 이화작용(2) N NH H O NH₂ 구아닌 H₂O NH4 + N NH H O O₂+H₂O H₂O₂ N NH H O
잔틴 산화효소 히포잔틴 잔틴

107
퓨린의 이화작용(3) H₂O+O₂ H₂O₂ HN NH N H O N NH H O 우르산 잔틴 HN NH N H O ¯O H+
산화효소 우르산 잔틴 HN NH N H O ¯O H+ 우르산염

108
누클레오티드 대사의 혼란에 의한 질병 ① 중증 합병형 면역 결핍증 (Severe Combined ImmunoDeficiency, SCID) ② 통풍(Gout) ③ 레쉬-니한 증후군(Lesch-Nyhan syndrome)
 ② 통풍(Gout) ③ 레쉬-니한 증후군(Lesch-Nyhan syndrome)")
109
(Severe Combined ImmunoDeficiency, SCID)
① 중증 합병형 면역 결핍증 (Severe Combined ImmunoDeficiency, SCID) 유발 원인 아데노신 탈아미노화효소 활성의 결핍 증상 반복적 감염 이른 나이에 사망 T세포 감소 치료법 조기 치료과정 : 환자를 환경으로부터 완전히 격리시킴 척수 이식 유전자 치료
 유발 원인. 아데노신 탈아미노화효소 활성의 결핍. 증상. 반복적 감염. 이른 나이에 사망. T세포 감소. 치료법. 조기 치료과정. : 환자를 환경으로부터 완전히 격리시킴. 척수 이식. 유전자 치료.")
110
② 통풍(Gout) 유발 원인 혈청 내에 존재하는 높은 농도의 우르산염 → 혈청 내 우르산의 증가
: 이른 나이에 신장 결석의 형성을 유발 ∴ 통풍 증상으로 이어짐 증상 고통스러운 관절염 감염 우르산염 결정의 축적에 의해 신장 손상

111
치료법 알로푸라놀(allopurinol)의 복용 히포잔틴과 잔틴으로부터 우르산염의 합성 감소 히포잔틴, 잔틴의 혈청 농도 증가
우르산염의 혈청 농도 감소 * 알로푸라놀(allopurinol) 통풍 치료약 히포잔틴의 유사체 → 처음: 기질로 작용 나중: 잔틴 산화효소의 억제제로 작용 복용 후
 통풍 치료약. 히포잔틴의 유사체. → 처음: 기질로 작용. 나중: 잔틴 산화효소의. 억제제로 작용. 복용 후.")
112
③ 레쉬-니한 증후군(Lesch-Nyhan syndrome)
유발 원인: 회수 경로의 돌연변이 구아닐산과 이노신산의 회수 경로에 작용하는 히포잔틴-구아닌 포스포리보실기 전이효소의 결핍 PRPP의 농도 증가, 퓨린 신생합성 속도의 증가, 우르산의 과대생성 증상 선천성 대사 이상: 강제성 자해 행위 어린이: 2~3살 때 손가락과 입술을 물어뜯음 (억제되지 않으면 손가락, 입술을 씹음) 공격적 행동→극단적인 적개심 정신지체 뇌성마비
 공격적 행동→극단적인 적개심. 정신지체. 뇌성마비.")
113
치료법 알로푸라놀을 투여하여 요산 생성을 억제하는 방법 고요산혈증에 의한 신부전, 통풍 등을 예방
신경 증상에 대한 근본적인 치료법이 아직까지는 없는 실정 행동치료를 통하여 자해를 예방 억제 요법이나 발치를 통하여 자해에 의한 손상을 감소 불안, 공격적 행동 등을 안정화시키기 위해서 신경안정제, 항경련제 등이 도움

114
Chapter 31 아미노산의 분해와 요소 회로

115
목 차 3. Chapter 31 아미노산의 분해와 요소 회로 31.1 아미노산 분해의 첫 단계는 질소의 제거이다.
목 차 3. Chapter 31 아미노산의 분해와 요소 회로 31.1 아미노산 분해의 첫 단계는 질소의 제거이다. 31.2 육지에 사는 대부분의 척추동물에서 암모늄 이온은 요소로 전환된다. 31.3 대사경로의 주요 중간산물들로 나타난다.

116
31.1 아미노산 분해의 첫 단계는 질소의 제거이다 1. 아미노산의 탈아미노 반응 ⑴ 산화적 탈아미노반응
31.1 아미노산 분해의 첫 단계는 질소의 제거이다 1. 아미노산의 탈아미노 반응 ⑴ 산화적 탈아미노반응 생체내 아미노산분해반응의 하나. RCH(NH2)COOH→RCOCOOH+NH3 아미노산을 적당한 수소 수용체로 산화해 α-케토산과 암모니아로 변화시키는 반응이며, 다음의 효소에 의해 촉매된다. 글루타민산 디하이드로제나아제 : NAD 또는 NADP를 보효소로 해서, 글루타민산을 탈아미노하는 작용이 있다. 아미노기 전이반응으로 글루타민산에 모여진 거의 모든 아미노산의 아미노기는 이 반응에 의해서 대사된다.
COOH→RCOCOOH+NH3. 아미노산을 적당한 수소 수용체로 산화해 α-케토산과 암모니아로 변화시키는 반응이며, 다음의 효소에 의해 촉매된다. 글루타민산 디하이드로제나아제 : NAD 또는 NADP를 보효소로 해서, 글루타민산을 탈아미노하는 작용이 있다. 아미노기 전이반응으로 글루타민산에 모여진 거의 모든 아미노산의 아미노기는 이 반응에 의해서 대사된다.")
117
⑵ 비산화적 탈아미노반응 아미노산에 pyridoxal phosphate를 보효소로 하는
탈수소효소(dehydratase)가 작용하면 비산화적 탈아미노반응이 생성 탈수소효소(dehydratase) - serine dehydratase - threonine dehydratase - cysteine desulfhydrolase(탈황화수소효소) methionine은 homocysteine → cystathionine을 거쳐 cysteine으로 됨
가 작용하면 비산화적. 탈아미노반응이 생성. 탈수소효소(dehydratase) - serine dehydratase. - threonine dehydratase. - cysteine desulfhydrolase(탈황화수소효소) methionine은 homocysteine → cystathionine을. 거쳐 cysteine으로 됨.")
118
31.2 육지에 사는 대부분의 척추동물에서 암모늄 이온은 요소로 전환된다
31.2 육지에 사는 대부분의 척추동물에서 암모늄 이온은 요소로 전환된다 탈아미노반응에서 생긴 암모니아는 자유롭게 mitochondria 막을 통과한다. 혈액 중의 전 암모니아 양의 1%는 NH3형으로 존재하고 나머지는 암모늄이온(NH4+)형으로 존재한다. 암모늄이온은 형질막이나 mitochondria막을 통과할 수 없다. 이러한 독성은 암모니아가 TCA 회로의 구성성분인 α-ketoglutarate를 이 계에서 제거하기 때문인 것으로 생각하고 있다. 뇌에서는 glutamine을 생성하여 암모니아를 간이나 신장으로 운반한다.
형으로 존재한다. 암모늄이온은 형질막이나 mitochondria막을 통과할 수 없다. 이러한 독성은 암모니아가 TCA 회로의 구성성분인 α-ketoglutarate를 이 계에서 제거하기 때문인 것으로 생각하고 있다. 뇌에서는 glutamine을 생성하여 암모니아를 간이나 신장으로 운반한다.")
119
요소회로 (1) 암모니아는 카바모일인산생성효소에 의해 카바모일인산을 생성한다. (2) 카바모일인산의 카르바모일기가 오르니틴으로 전이되어 시트룰린을 생성한다. (3) 시트룰린이 아스파르트산과 반응하여 아르기노숙신산을 생성한다. (4) 아르기니노숙신산이 아르기닌과 푸마르산으로 분해된다. (5) 아르기닌이가수분해되어 요소를 형성하거나 오르니틴이 재생된다.
 암모니아는 카바모일인산생성효소에 의해 카바모일인산을 생성한다. (2) 카바모일인산의 카르바모일기가 오르니틴으로 전이되어 시트룰린을 생성한다. (3) 시트룰린이 아스파르트산과 반응하여 아르기노숙신산을 생성한다. (4) 아르기니노숙신산이 아르기닌과 푸마르산으로 분해된다. (5) 아르기닌이가수분해되어 요소를 형성하거나 오르니틴이 재생된다.")
120
31.3 분해된 아미노산의 탄소 원자들은 대사경로의 주요 중간산물들로 나타난다
31.3 분해된 아미노산의 탄소 원자들은 대사경로의 주요 중간산물들로 나타난다 아미노산 분해된 후의 탄소 골격들은 글루코오스로 전환 되거나 시트르산 회로로 들어갈 수 있다.

121
1. 피루브산은 많은 아미노산 대사의 진입점이다. 알라닌, 트립토판, 글리신, 세린, 시스테인, 트레오닌,
1. 피루브산은 많은 아미노산 대사의 진입점이다. 알라닌, 트립토판, 글리신, 세린, 시스테인, 트레오닌, 아미노기 이동으로 피루브산이 될 수 있다.

122
2. 옥살로아세트산은 대사 경로의 또 다른 진입점이다
아스파르트산과 아스파라긴은 아미노기 이동반응으로 옥살로아세트산이 된다. 아스팔트산의 아미노기는 옥살로 아세트산에 전달된다. 아스파라긴은 아스파라긴가수분해 효소에 의해 암모늄이온과 아스팔트산으로 가수분해 된다. 아스팔트산+a-ketoglutarate 옥살로아세트산+글루탐산

123
3. α-케토글루타르산은 오탄소 아미노산의 진입점이다
3. α-케토글루타르산은 오탄소 아미노산의 진입점이다 히스티딘, 글루타민, 프롤린, 아르기닌, 글루탐산 등의 오탄소의 아미노산들은 α–케토글루타르산으로 바뀐다.

124
히스티딘 분해 히스티딘은 중간산물인 4-이미다졸론 5-프로피온산을 통해 글루탐산으로 전환
중간산물의 고리에 아미드결합은 글루탐산의 N-포름이미노 유도체로 가수분해, 글루탐산으로 전환

125
글루탐산 형성 글루타민은 글루타민 가수분해효소에 의해 글루탐산과 NH4+로 가수분해된다.
프롤린과 아르기닌은 각각 글루탐산 감마-세미알데히드로 전환되고 그 다음 글루탐산으로 산화된다.

126
4. 석시닐 CoA는 여러 무극성 아미노산들의 진입점이다
프로피오닐coA와 그 이후 메틸말로닐coA는 세가지 아미노산의 분해에서 생기는 중간산물이다.

127
5. 방향족 아미노산들의 분해에는 산소화효소가 필요하다
방향족 아미노산의 분해에서의 최종산물의 분해과정은 앞에서 나온 아미노산들처럼 간단하지가 않다. * 방향족 고리를 분해하기 위하여 산소 분자를 사용한다. - 산소화효소(oxygenase) * 생물학적으로 생체에 있는 방향족 고리들의 쪼개짐은 거의 다 이산소화효소(dioxygenase)들로 촉매된다.
 * 생물학적으로 생체에 있는 방향족 고리들의 쪼개짐은 거의 다 이산소화효소(dioxygenase)들로 촉매된다.")
128
지금까지 발표를 경청해 주셔서 감사합니다♥


? 초신성 폭발 원소들은.>")









 누가 앙금을 만들었는지 쉽게 알려 줘! 앙금 생성 반응식.>")

 생태계에서의 에너지 흐름 1. 에너지, 효소와 조절>")
는 변화를 의미. 세포의 모든 화학반응과 물리적인 일에 관련되어 있다.>")




 유전학적으로 본 병인의 분류>")
